

Motor number controls cargo switching at actin-microtubule intersections in vitro
- PMID: 20399098
- PMCID: PMC2934746
- DOI: 10.1016/j.cub.2010.03.024
Motor number controls cargo switching at actin-microtubule intersections in vitro
Abstract
Background: Cellular activities such as endocytosis and secretion require that cargos actively switch between the microtubule (MT) and actin filament (AF) networks. Cellular studies suggest that switching may involve a tug of war or coordinate regulation of MT- and AF-based motor function.
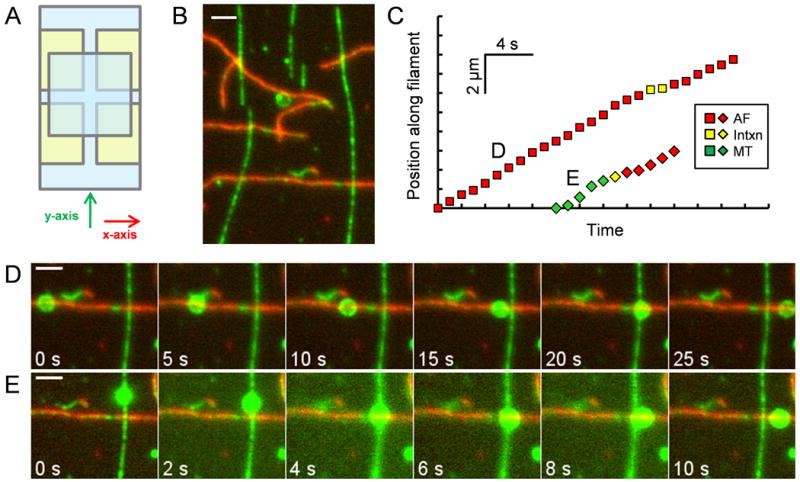
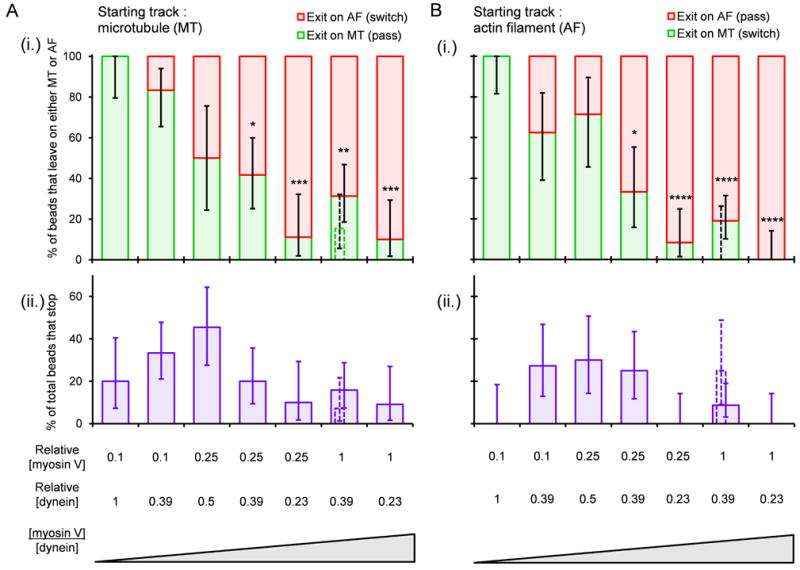
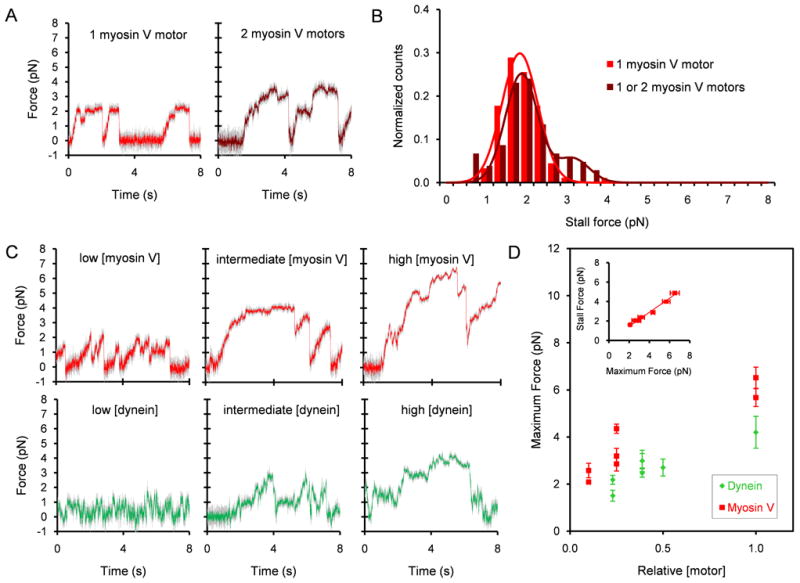
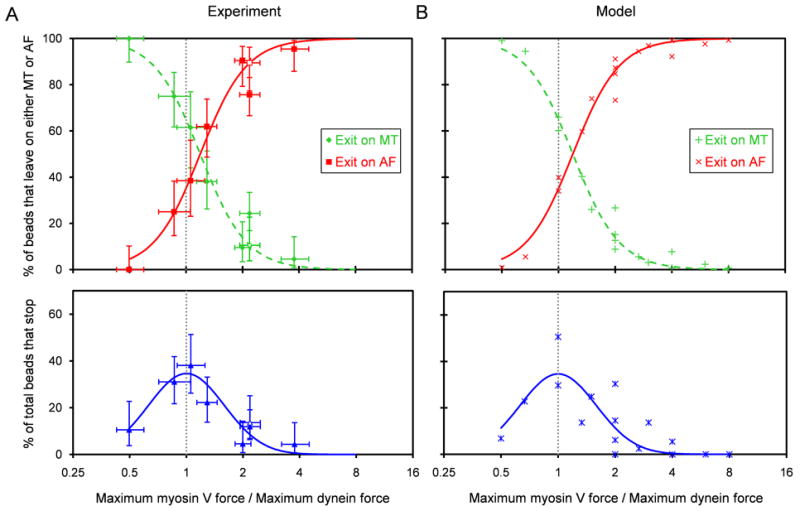
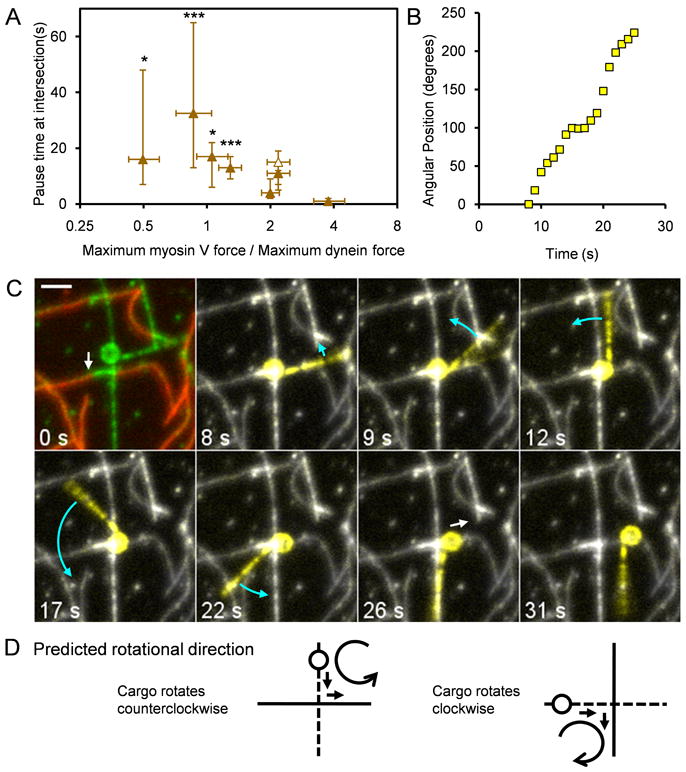
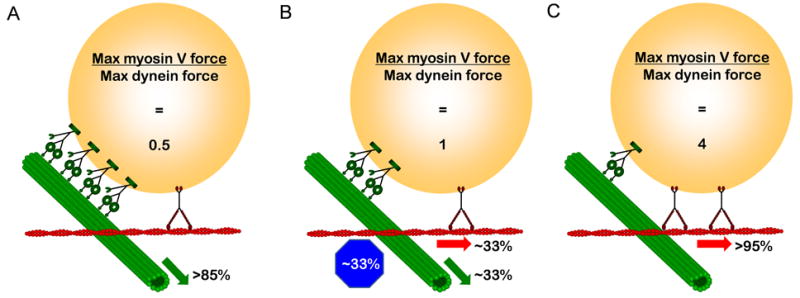
Results: To test the hypothesis that motor number can be used to direct the outcome of a tug-of-war process, we reconstituted cargo switching at MT-AF intersections in a minimal system with purified myosin V and dynein-dynactin motors bound to beads. Beads containing both motors often paused at the intersections and rotated about an axis perpendicular to both filaments, suggesting that competing motors apply a torque on their cargo. Force measurements showed that motor forces scale with the number of engaged myosin V and dynein-dynactin motors. Whether beads remained on a MT or AF or switched to the alternate track was determined by which set of motors collectively produced greater force. Passing and switching probabilities were similar whether the bead approached an intersection on either a MT or an AF. Beads with a force ratio near unity had approximately equal probabilities of exiting on the MT, exiting on the AF, or remaining stalled at the intersection. A simple statistical model quantitatively describes the relationship between switching probability and motor number.
Conclusions: Cargo switching can be tuned via combinations of 1-4 myosin V and 1-4 dynein-dynactin engaged motors through a simple force-mediated mechanism.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Caviston JP, Holzbaur EL. Microtubule motors at the intersection of trafficking and transport. Trends Cell Biol. 2006;16:530–537. - PubMed
-
- Guzik BW, Goldstein LS. Microtubule-dependent transport in neurons: steps towards an understanding of regulation, function and dysfunction. Curr Opin Cell Biol. 2004;16:443–450. - PubMed
-
- Vale RD. The molecular motor toolbox for intracellular transport. Cell. 2003;112:467–480. - PubMed
-
- Krendel M, Mooseker MS. Myosins: tails (and heads) of functional diversity. Physiology (Bethesda) 2005;20:239–251. - PubMed
-
- Kuznetsov SA, Langford GM, Weiss DG. Actin-dependent organelle movement in squid axoplasm. Nature. 1992;356:722–725. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
