

Zebrafish MTMR14 is required for excitation-contraction coupling, developmental motor function and the regulation of autophagy
- PMID: 20400459
- PMCID: PMC2883342
- DOI: 10.1093/hmg/ddq153
Zebrafish MTMR14 is required for excitation-contraction coupling, developmental motor function and the regulation of autophagy
Abstract
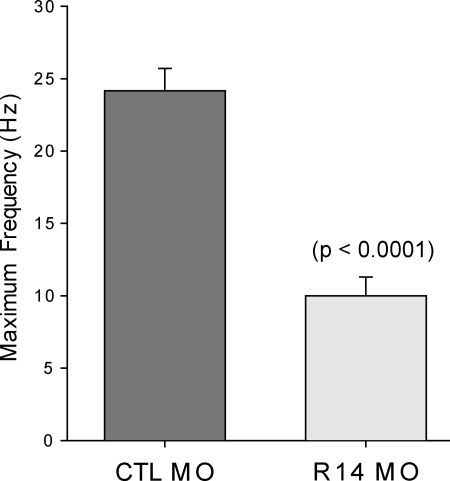
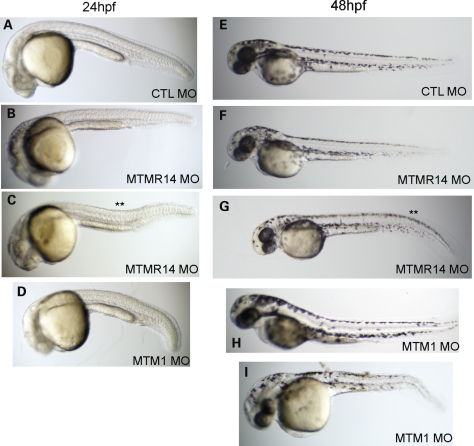
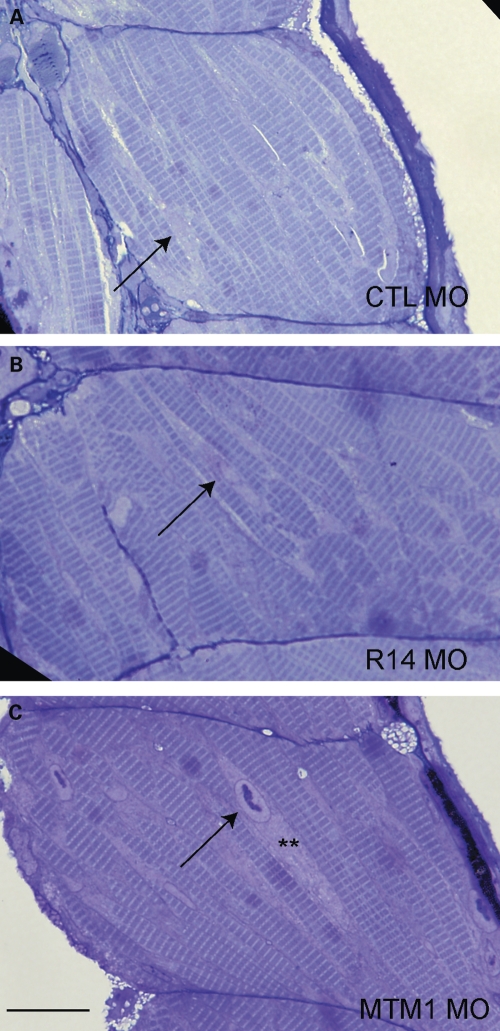
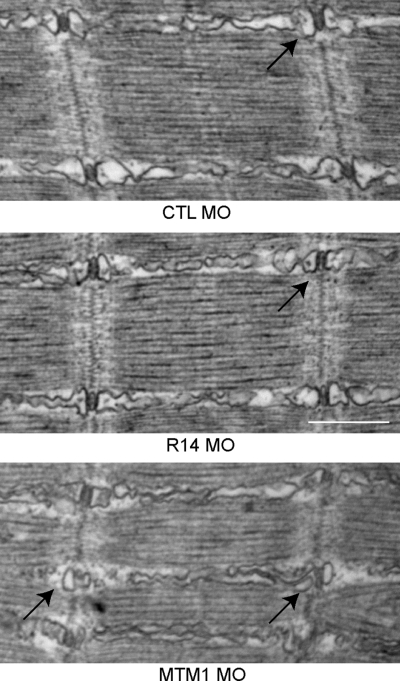
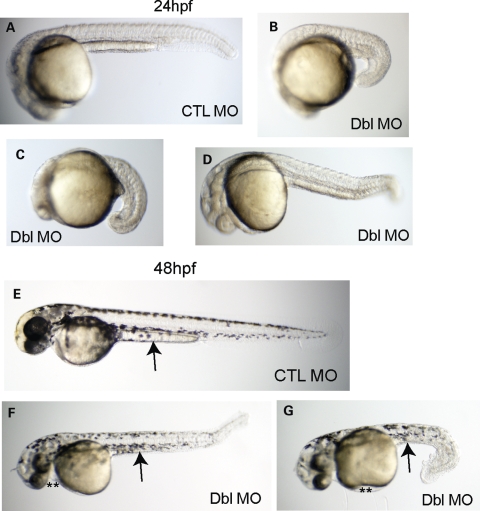
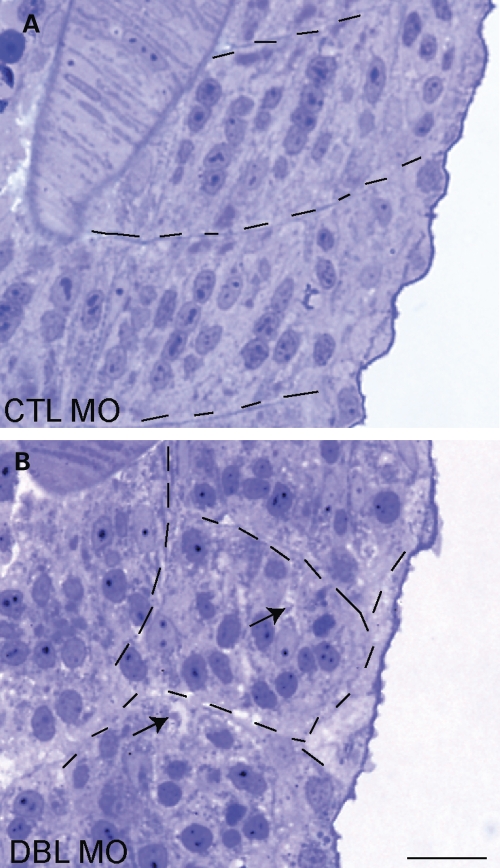
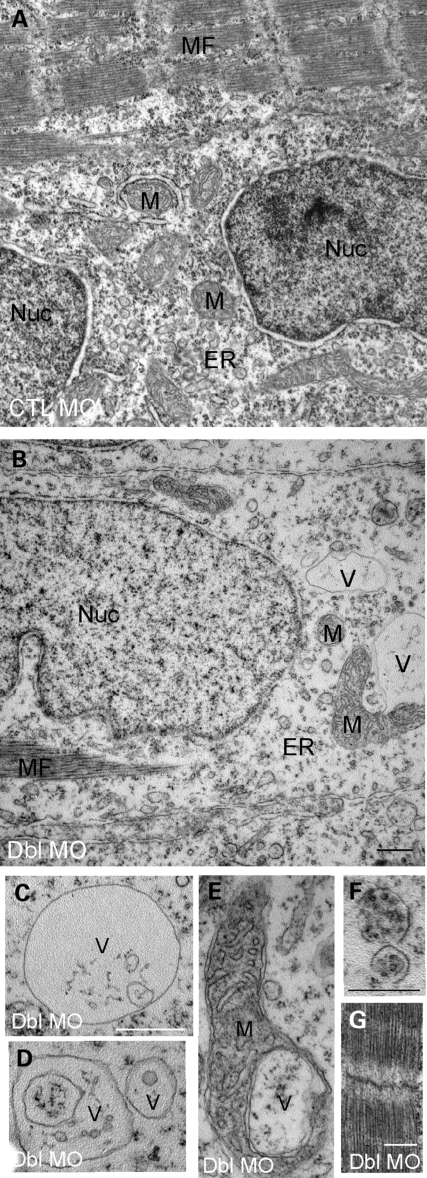
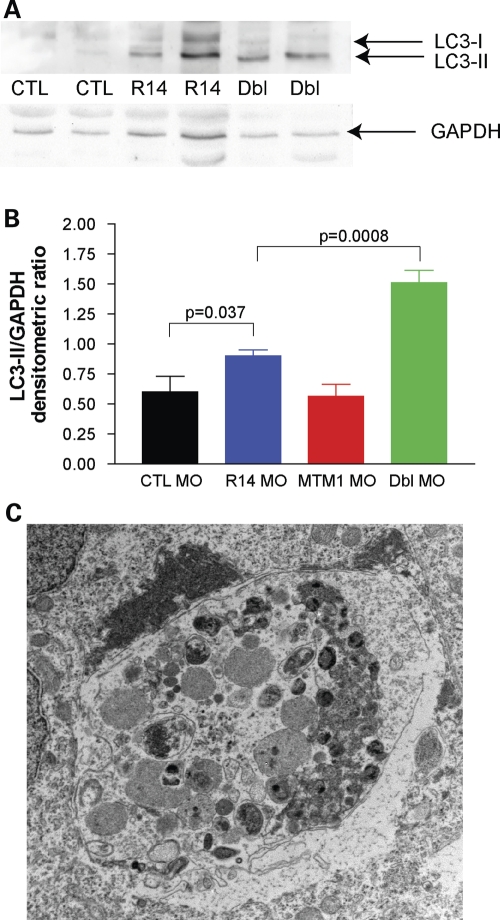
Myotubularins are a family of dual-specificity phosphatases that act to modify phosphoinositides and regulate membrane traffic. Mutations in several myotubularins are associated with human disease. Sequence changes in MTM1 and MTMR14 (also known as Jumpy) have been detected in patients with a severe skeletal myopathy called centronuclear myopathy. MTM1 has been characterized in vitro and in several model systems, while the function of MTMR14 and its specific role in muscle development and disease is much less well understood. We have previously reported that knockdown of zebrafish MTM1 results in significantly impaired motor function and severe histopathologic changes in skeletal muscle that are characteristic of human centronuclear myopathy. In the current study, we examine zebrafish MTMR14 using gene dosage manipulation. As with MTM1 knockdown, morpholino-mediated knockdown of MTMR14 results in morphologic abnormalities, a developmental motor phenotype characterized by diminished spontaneous contractions and abnormal escape response, and impaired excitation-contraction coupling. In contrast to MTM1 knockdown, however, muscle ultrastructure is unaffected. Double knockdown of both MTM1 and MTMR14 significantly impairs motor function and alters skeletal muscle ultrastructure. The combined effect of reducing levels of both MTMR14 and MTM1 is significantly more severe than either knockdown alone, an effect which is likely mediated, at least in part, by increased autophagy. In all, our results suggest that MTMR14 is required for motor function and, in combination with MTM1, is required for myocyte homeostasis and normal embryonic development.
Figures

References
-
- Miaczynska M., Stenmark H. Mechanisms and functions of endocytosis. J. Cell Biol. 2008;180:7–11. doi:10.1083/jcb.200711073. - DOI - PMC - PubMed
-
- Di Paolo G., De Camilli P. Phosphoinositides in cell regulation and membrane dynamics. Nature. 2006;443:651–657. doi:10.1038/nature05185. - DOI - PubMed
-
- Robinson F.L., Dixon J.E. Myotubularin phosphatases: policing 3-phosphoinositides. Trends Cell Biol. 2006;16:403–412. doi:10.1016/j.tcb.2006.06.001. - DOI - PubMed
-
- De Matteis M.A., Godi A. PI-loting membrane traffic. Nat. Cell Biol. 2004;6:487–492. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
