

Unpaired 5' ppp-nucleotides, as found in arenavirus double-stranded RNA panhandles, are not recognized by RIG-I
- PMID: 20400512
- PMCID: PMC2881745
- DOI: 10.1074/jbc.M109.089425
Unpaired 5' ppp-nucleotides, as found in arenavirus double-stranded RNA panhandles, are not recognized by RIG-I
Abstract
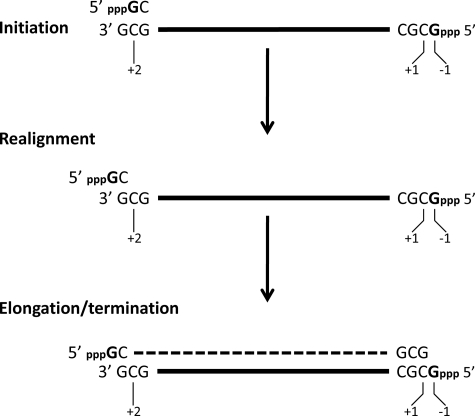
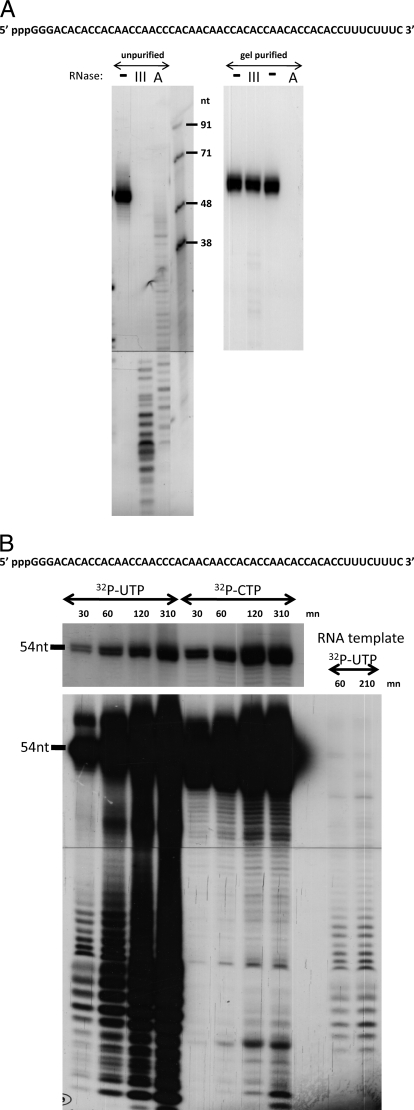
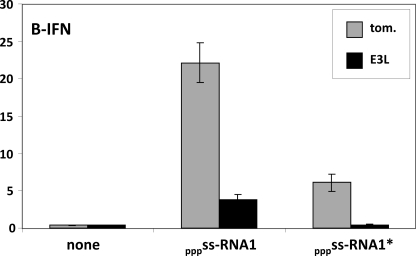
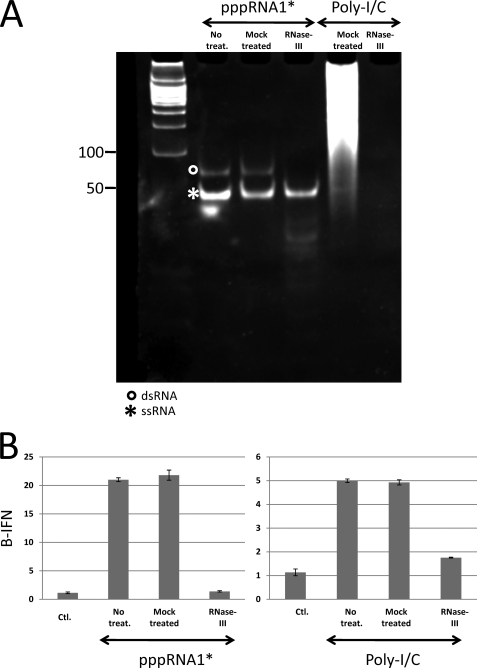
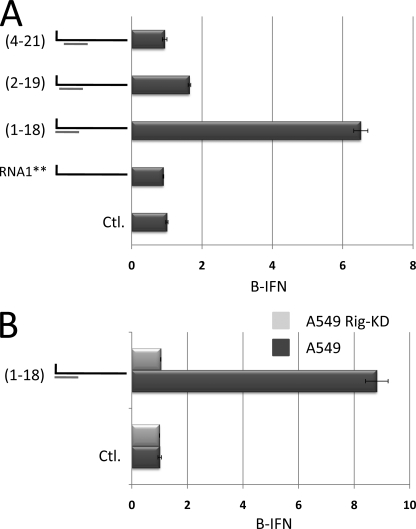
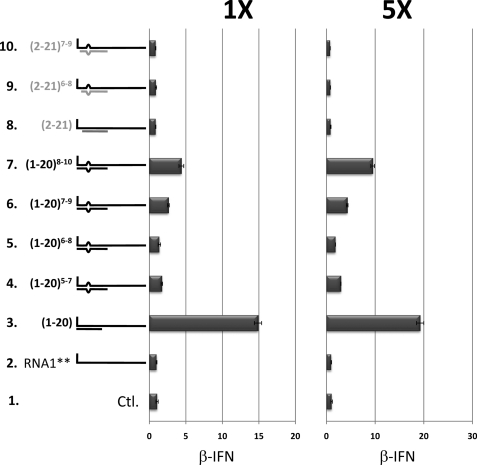
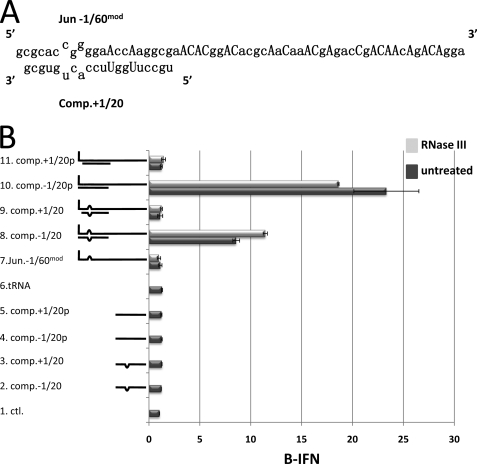
Arenavirus and bunyavirus RNA genomes are unusual in that they are found in circular nucleocapsids, presumably due to the annealing of their complementary terminal sequences. Moreover, arenavirus genome synthesis initiates with GTP at position +2 of the template rather than at the precise 3' end (position +1). After formation of a dinucleotide, 5' pppGpC(OH) is then realigned on the template before this primer is extended. The net result of this "prime and realign" mechanism of genome initiation is that 5' pppG is found as an unpaired 5' nucleotide when the complementary genome ends anneal to form a double-stranded (dsRNA) panhandle. Using 5' pppRNA made in vitro and purified so that all dsRNA side products are absent, we have determined that both this 5' nucleotide overhang, as well as mismatches within the dsRNA (as found in some arenavirus genomes), clearly reduce the ability of these model dsRNAs to induce interferon upon transfection into cells. The presence of this unpaired 5' ppp-nucleotide is thus another way that some viruses appear to use to avoid detection by cytoplasmic pattern recognition receptors.
Figures
Similar articles
-
Short double-stranded RNAs with an overhanging 5' ppp-nucleotide, as found in arenavirus genomes, act as RIG-I decoys.J Biol Chem. 2011 Feb 25;286(8):6108-16. doi: 10.1074/jbc.M110.186262. Epub 2010 Dec 15. J Biol Chem. 2011. PMID: 21159780 Free PMC article.
-
Visualization of Double-Stranded RNA Colocalizing With Pattern Recognition Receptors in Arenavirus Infected Cells.Front Cell Infect Microbiol. 2018 Jul 24;8:251. doi: 10.3389/fcimb.2018.00251. eCollection 2018. Front Cell Infect Microbiol. 2018. PMID: 30087859 Free PMC article.
-
Alternative translation contributes to the generation of a cytoplasmic subpopulation of the Junín virus nucleoprotein that inhibits caspase activation and innate immunity.J Virol. 2024 Feb 20;98(2):e0197523. doi: 10.1128/jvi.01975-23. Epub 2024 Jan 31. J Virol. 2024. PMID: 38294249 Free PMC article.
-
[Viral sensing by RNA helicases].Virologie (Montrouge). 2020 Dec 1;24(6):419-436. doi: 10.1684/vir.2020.0871. Virologie (Montrouge). 2020. PMID: 33441291 Review. French.
-
Viral sensing by RNA helicases.Virologie (Montrouge). 2020 Dec 1;24(6):36-52. doi: 10.1684/vir.2020.0872. Virologie (Montrouge). 2020. PMID: 33441288 Review.
Cited by
-
RIG-I Detects Kaposi's Sarcoma-Associated Herpesvirus Transcripts in a RNA Polymerase III-Independent Manner.mBio. 2018 Jul 3;9(4):e00823-18. doi: 10.1128/mBio.00823-18. mBio. 2018. PMID: 29970461 Free PMC article.
-
Environmentally Triggerable Retinoic Acid-Inducible Gene I Agonists Using Synthetic Polymer Overhangs.Bioconjug Chem. 2018 Mar 21;29(3):742-747. doi: 10.1021/acs.bioconjchem.7b00697. Epub 2018 Jan 19. Bioconjug Chem. 2018. PMID: 29350913 Free PMC article.
-
Regulation of RIG-I-like receptor-mediated signaling: interaction between host and viral factors.Cell Mol Immunol. 2021 Mar;18(3):539-555. doi: 10.1038/s41423-020-00602-7. Epub 2021 Jan 18. Cell Mol Immunol. 2021. PMID: 33462384 Free PMC article. Review.
-
Short double-stranded RNAs with an overhanging 5' ppp-nucleotide, as found in arenavirus genomes, act as RIG-I decoys.J Biol Chem. 2011 Feb 25;286(8):6108-16. doi: 10.1074/jbc.M110.186262. Epub 2010 Dec 15. J Biol Chem. 2011. PMID: 21159780 Free PMC article.
-
Mismatches in the Influenza A Virus RNA Panhandle Prevent Retinoic Acid-Inducible Gene I (RIG-I) Sensing by Impairing RNA/RIG-I Complex Formation.J Virol. 2015 Oct 7;90(1):586-90. doi: 10.1128/JVI.01671-15. Print 2016 Jan 1. J Virol. 2015. PMID: 26446607 Free PMC article.
References
-
- Prehaud C., Lopez N., Blok M. J., Obry V., Bouloy M. (1997) Virology 227, 189–197 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
