

RNA-dependent RNA polymerase 1 from Nicotiana tabacum suppresses RNA silencing and enhances viral infection in Nicotiana benthamiana
- PMID: 20400679
- PMCID: PMC2879737
- DOI: 10.1105/tpc.109.072058
RNA-dependent RNA polymerase 1 from Nicotiana tabacum suppresses RNA silencing and enhances viral infection in Nicotiana benthamiana
Abstract
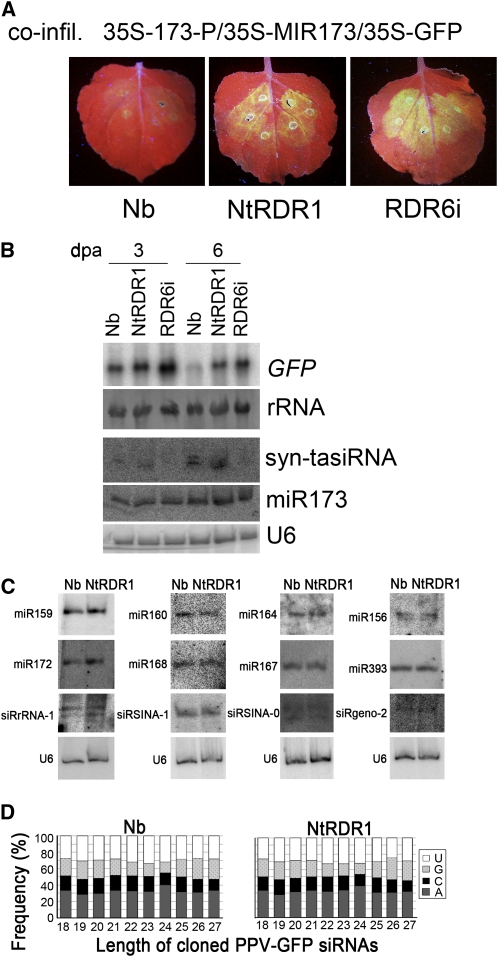
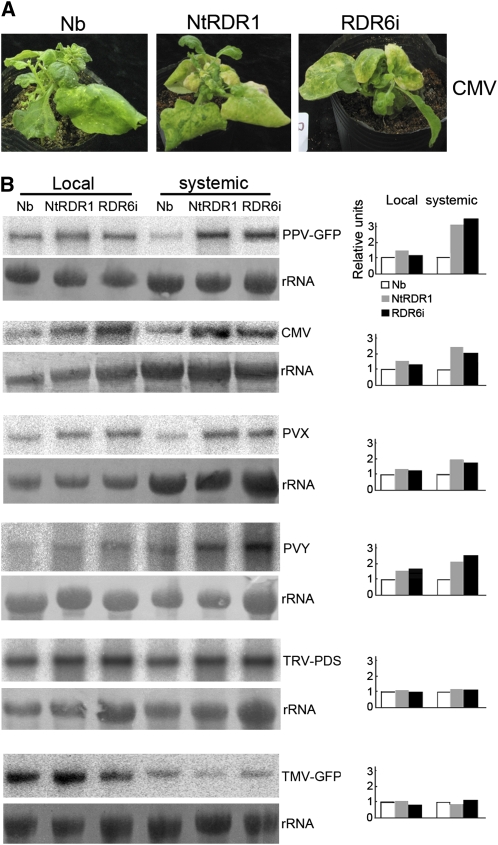
Endogenous eukaryotic RNA-dependent RNA polymerases (RDRs) produce double-stranded RNA intermediates in diverse processes of small RNA synthesis in RNA silencing pathways. RDR6 is required in plants for posttranscriptional gene silencing induced by sense transgenes (S-PTGS) and has an important role in amplification of antiviral silencing. Whereas RDR1 is also involved in antiviral defense in plants, this does not necessarily proceed through triggering silencing. In this study, we show that Nicotiana benthamiana transformed with RDR1 from Nicotiana tabacum (Nt-RDR1 plants) exhibits hypersusceptibility to Plum pox potyvirus and other viruses, resembling RDR6-silenced (RDR6i) N. benthamiana. Analysis of transient induction of RNA silencing in N. benthamiana Nt-RDR1 and RDR6i plants revealed that Nt-RDR1 possesses silencing suppression activity. We found that Nt-RDR1 does not interfere with RDR6-dependent siRNA accumulation but turns out to suppress RDR6-dependent S-PTGS. Our results, together with previously published data, suggest that RDR1 might have a dual role, contributing, on one hand, to salicylic acid-mediated antiviral defense, and suppressing, on the other hand, the RDR6-mediated antiviral RNA silencing. We propose a scenario in which the natural loss-of-function variant of RDR1 in N. benthamiana may be the outcome of selective pressure to maintain a high RDR6-dependent antiviral defense, which would be required to face the hypersensitivity of this plant to a large number of viruses.
Figures
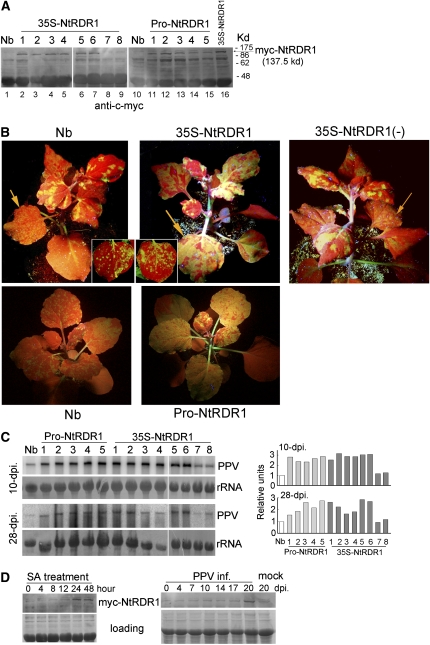
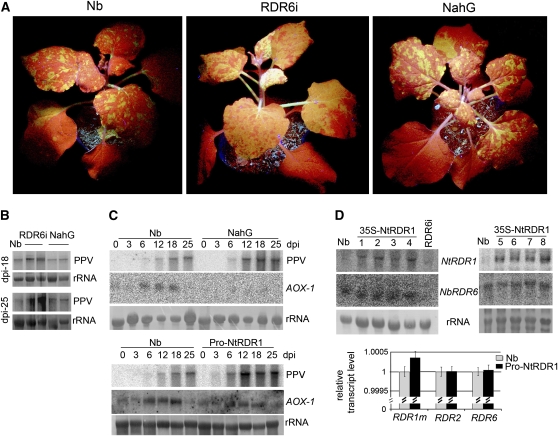
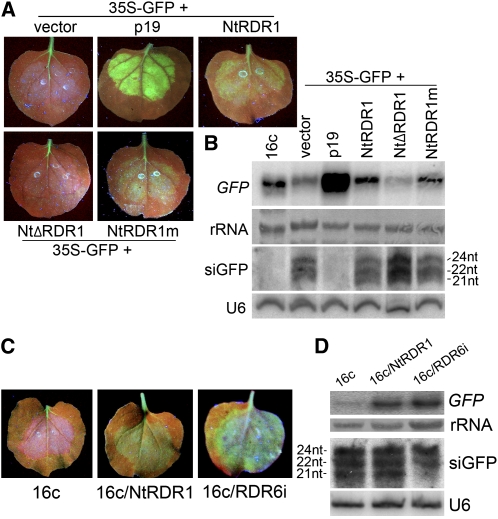
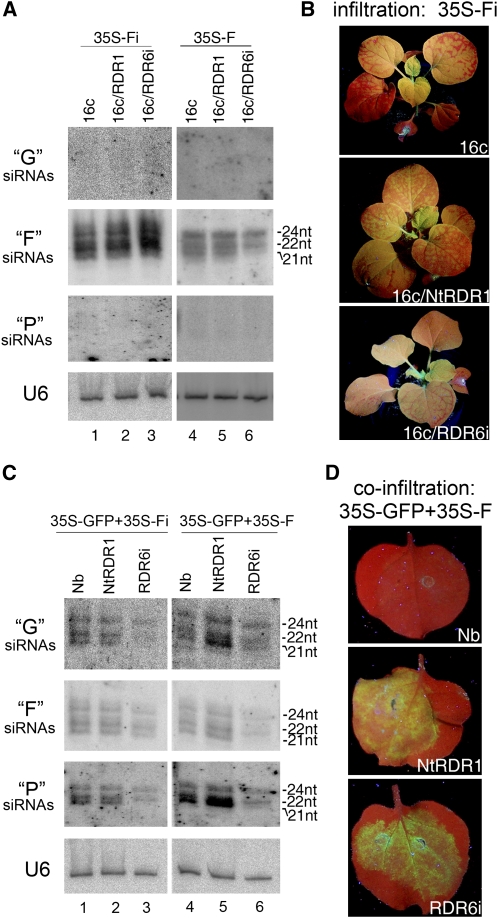
Similar articles
-
Tobacco RNA-dependent RNA polymerase 1 affects the expression of defence-related genes in Nicotiana benthamiana upon Tomato leaf curl Gujarat virus infection.Planta. 2020 Jul 1;252(1):11. doi: 10.1007/s00425-020-03417-y. Planta. 2020. PMID: 32613448
-
RNA-dependent RNA polymerase 6 delays accumulation and precludes meristem invasion of a viroid that replicates in the nucleus.J Virol. 2010 Mar;84(5):2477-89. doi: 10.1128/JVI.02336-09. Epub 2009 Dec 16. J Virol. 2010. PMID: 20015979 Free PMC article.
-
RNA-dependent RNA polymerase 1 in potato (Solanum tuberosum) and its relationship to other plant RNA-dependent RNA polymerases.Sci Rep. 2016 Mar 16;6:23082. doi: 10.1038/srep23082. Sci Rep. 2016. PMID: 26979928 Free PMC article.
-
Antiviral role of plant-encoded RNA-dependent RNA polymerases revisited with deep sequencing of small interfering RNAs of virus origin.Mol Plant Microbe Interact. 2010 Oct;23(10):1248-52. doi: 10.1094/MPMI-06-10-0124. Mol Plant Microbe Interact. 2010. PMID: 20831405 Review.
-
Recent advances in plant-virus interaction with emphasis on small interfering RNAs (siRNAs).Mol Biotechnol. 2013 Sep;55(1):63-77. doi: 10.1007/s12033-012-9615-7. Mol Biotechnol. 2013. PMID: 23086491 Review.
Cited by
-
Regulation of RNA-dependent RNA polymerase 1 and isochorismate synthase gene expression in Arabidopsis.PLoS One. 2013 Jun 17;8(6):e66530. doi: 10.1371/journal.pone.0066530. Print 2013. PLoS One. 2013. PMID: 23799112 Free PMC article.
-
Deficiency of multiple RNA silencing-associated genes may contribute to the increased susceptibility of Nicotiana benthamiana to viruses.Plant Cell Rep. 2024 Jun 19;43(7):177. doi: 10.1007/s00299-024-03262-3. Plant Cell Rep. 2024. PMID: 38898307 Free PMC article.
-
Involvement of RDR6 in short-range intercellular RNA silencing in Nicotiana benthamiana.Sci Rep. 2012;2:467. doi: 10.1038/srep00467. Epub 2012 Jun 22. Sci Rep. 2012. PMID: 22737403 Free PMC article.
-
Satellite RNA reduces expression of the 2b suppressor protein resulting in the attenuation of symptoms caused by Cucumber mosaic virus infection.Mol Plant Pathol. 2011 Aug;12(6):595-605. doi: 10.1111/j.1364-3703.2010.00696.x. Epub 2011 Jan 17. Mol Plant Pathol. 2011. PMID: 21722297 Free PMC article.
-
Validation of reference genes for gene expression studies in virus-infected Nicotiana benthamiana using quantitative real-time PCR.PLoS One. 2012;7(9):e46451. doi: 10.1371/journal.pone.0046451. Epub 2012 Sep 28. PLoS One. 2012. PMID: 23029521 Free PMC article.
References
-
- Abbink E.M., Tjernberg P.A., Bol J.F., Linthors J.M. (1998). Tobacco mosaic virus helicase domain induces necrosis in N gene-carrying tobacco in the absence of virus replication. Mol. Plant Microbe Interact. 11: 1242–1246
-
- Alamillo J.M., Saenz P., Garcia J.A. (2006). Salicylic acid-mediated and RNA-silencing defense mechanisms cooperate in the restriction of systemic spread of plum pox virus in tobacco. Plant J. 48: 217–227 - PubMed
-
- Allen E., Xie Z., Gustafson A.M., Carrington J.C. (2005). MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121: 207–221 - PubMed
-
- Asikainen S., Storvik M., Lakso M., Wong G. (2007). Whole genome microarray analysis of C. elegans rrf-3 and eri-1 mutants. FEBS Lett. 581: 5050–5054 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
