

LIS1 and NudE induce a persistent dynein force-producing state
- PMID: 20403325
- PMCID: PMC2881166
- DOI: 10.1016/j.cell.2010.02.035
LIS1 and NudE induce a persistent dynein force-producing state
Abstract
Cytoplasmic dynein is responsible for many aspects of cellular and subcellular movement. LIS1, NudE, and NudEL are dynein interactors initially implicated in brain developmental disease but now known to be required in cell migration, nuclear, centrosomal, and microtubule transport, mitosis, and growth cone motility. Identification of a specific role for these proteins in cytoplasmic dynein motor regulation has remained elusive. We find that NudE stably recruits LIS1 to the dynein holoenzyme molecule, where LIS1 interacts with the motor domain during the prepowerstroke state of the dynein crossbridge cycle. NudE abrogates dynein force production, whereas LIS1 alone or with NudE induces a persistent-force dynein state that improves ensemble function of multiple dyneins for transport under high-load conditions. These results likely explain the requirement for LIS1 and NudE in the transport of nuclei, centrosomes, chromosomes, and the microtubule cytoskeleton as well as the particular sensitivity of migrating neurons to reduced LIS1 expression.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
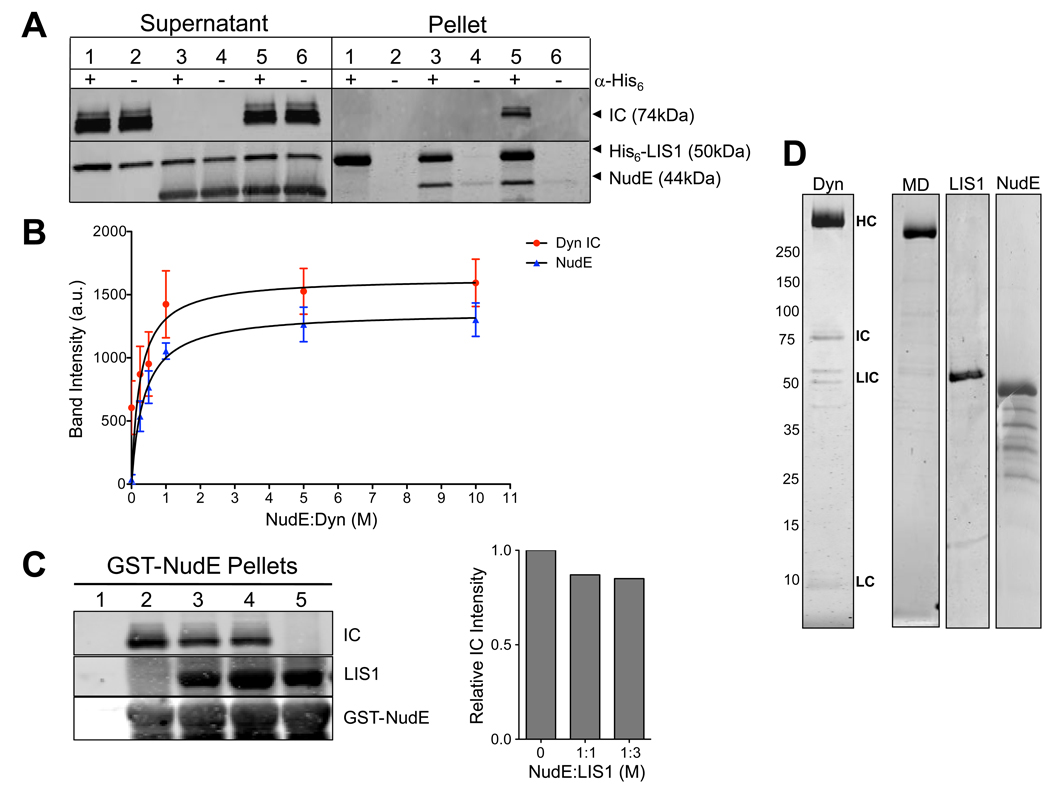
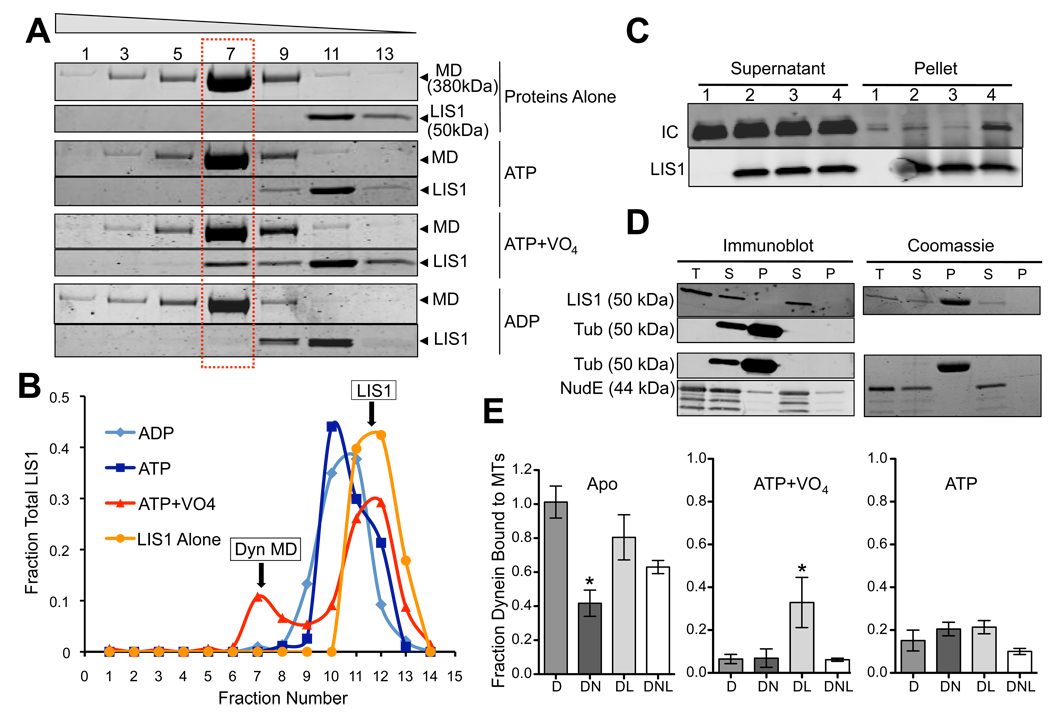
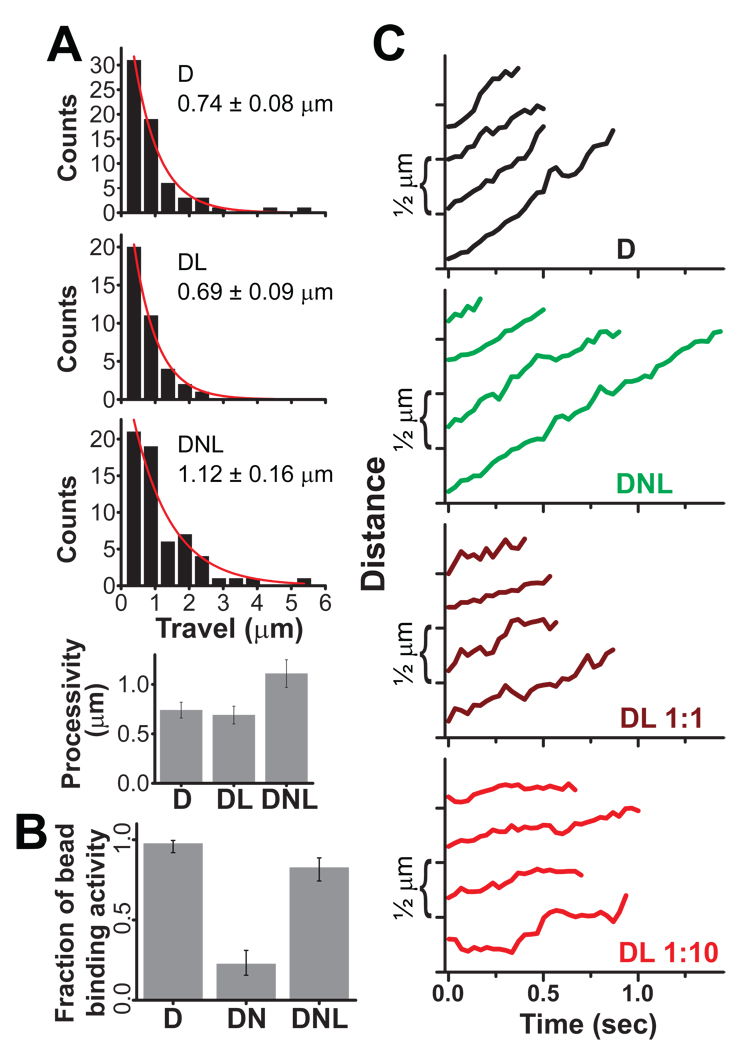
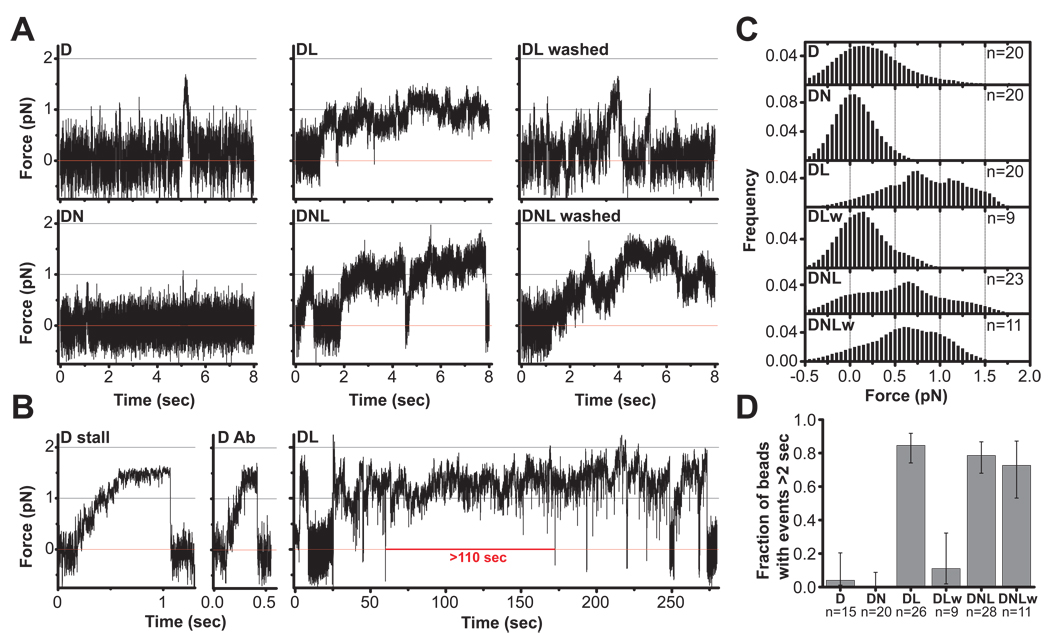
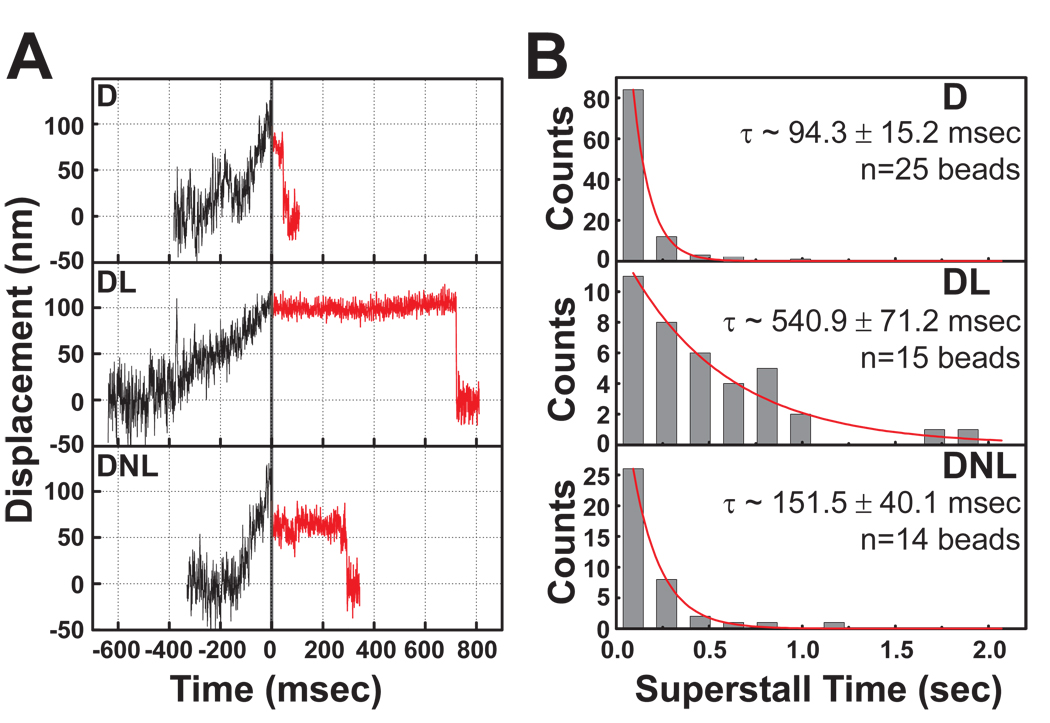
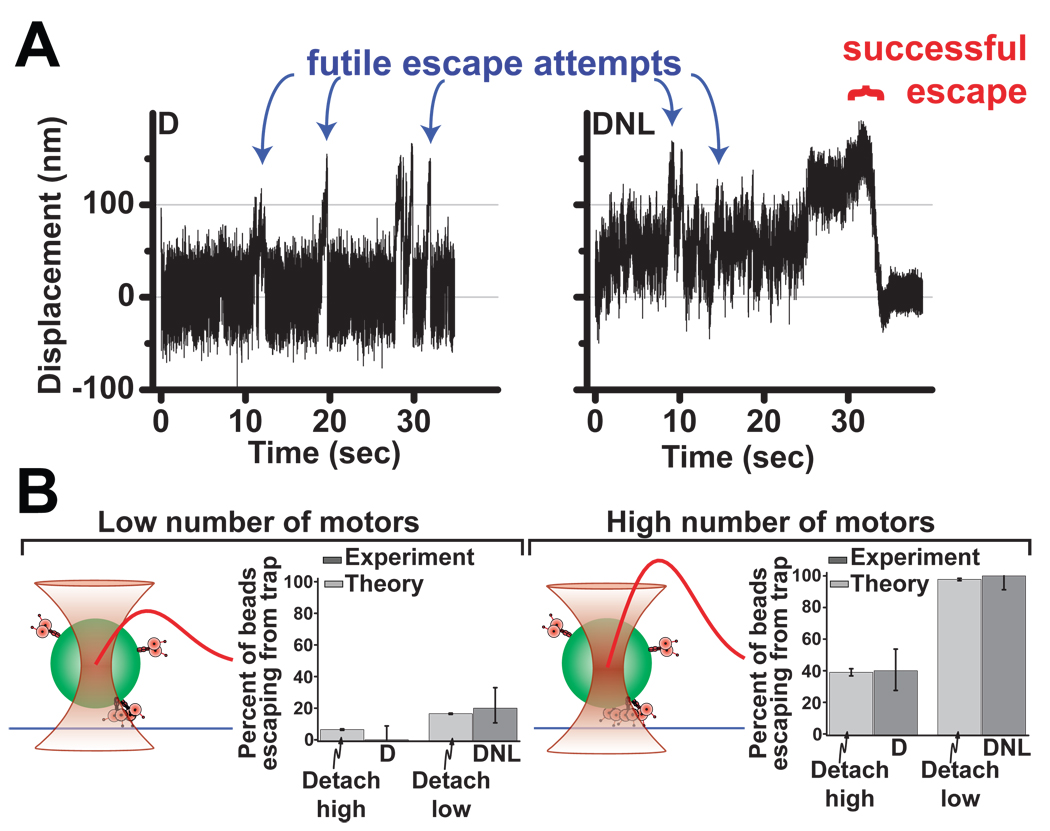
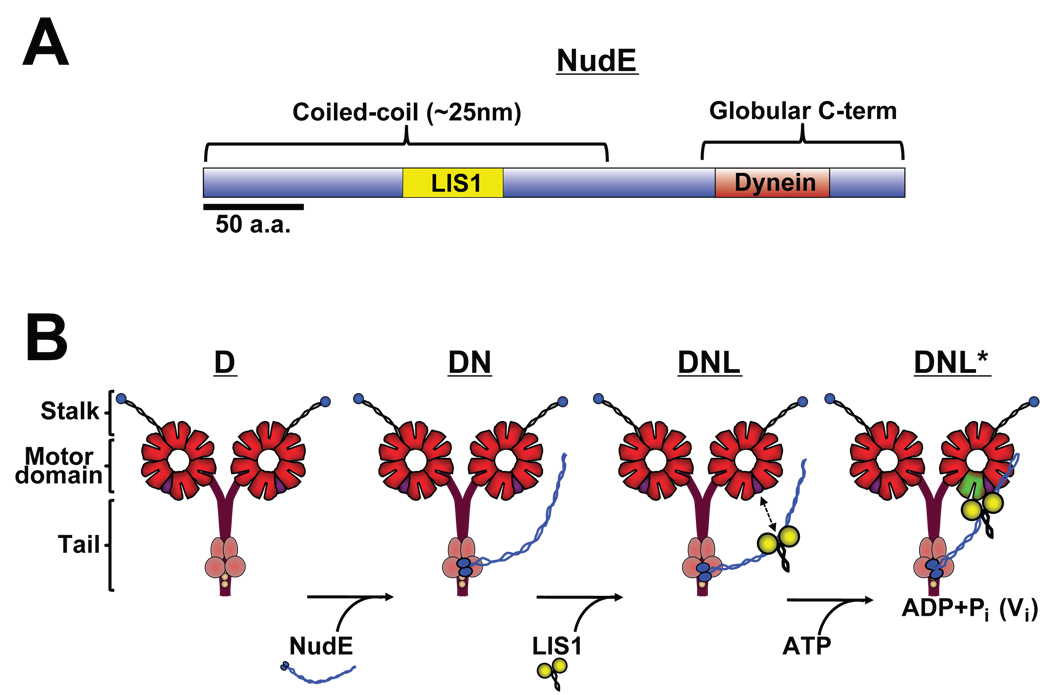
References
-
- Burgess SA, Walker ML, Sakakibara H, Knight PJ, Oiwa K. Dynein structure and power stroke. Nature. 2003;421:715–718. - PubMed
-
- Derewenda U, Tarricone C, Choi WC, Cooper DR, Lukasik S, Perrina F, Tripathy A, Kim MH, Cafiso DS, Musacchio A, Derewenda ZS. The structure of the coiled-coil domain of Ndel1 and the basis of its interaction with Lis1, the causal protein of Miller-Dieker lissencephaly. Structure. 2007;15:1467–1481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
