

The Hsp90 co-chaperone p23 of Toxoplasma gondii: Identification, functional analysis and dynamic interactome determination
- PMID: 20403389
- PMCID: PMC2990952
- DOI: 10.1016/j.molbiopara.2010.04.004
The Hsp90 co-chaperone p23 of Toxoplasma gondii: Identification, functional analysis and dynamic interactome determination
Abstract
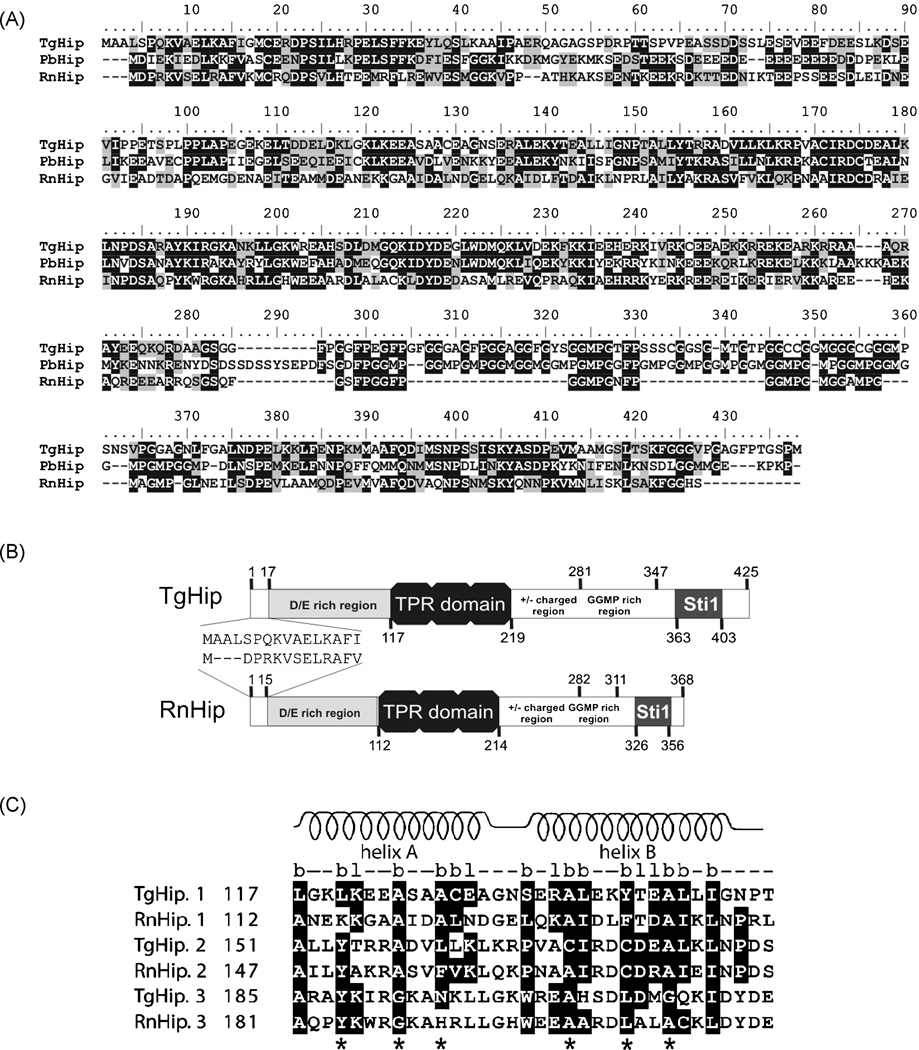
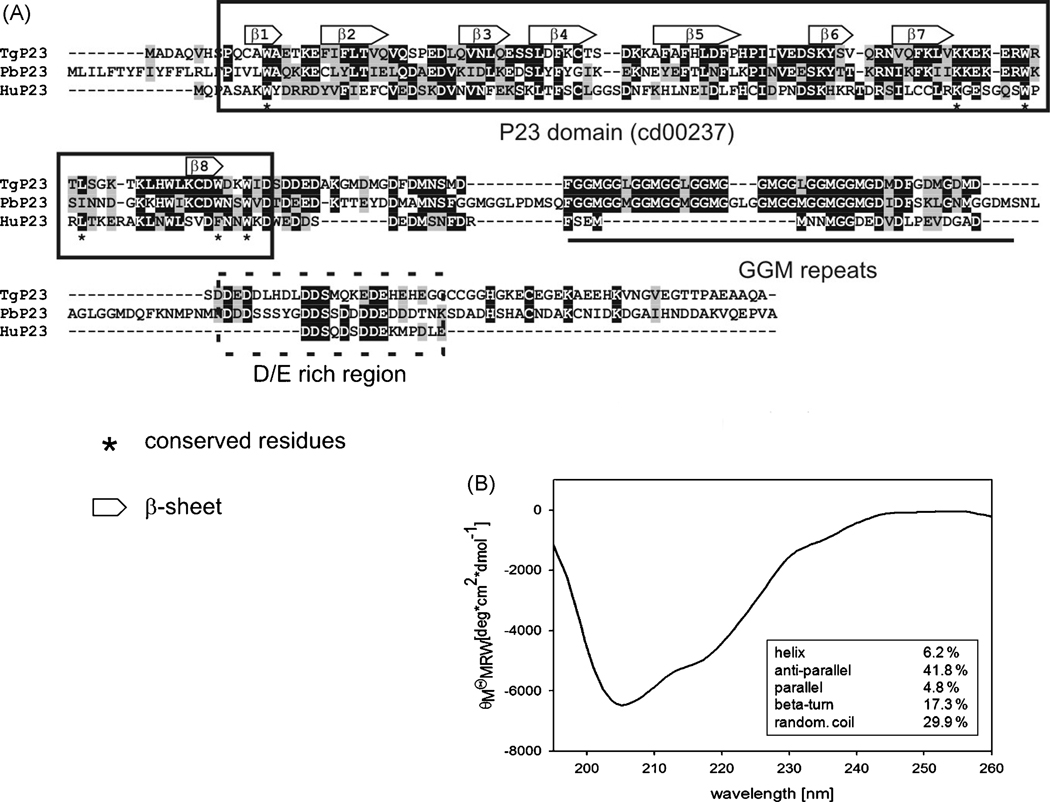
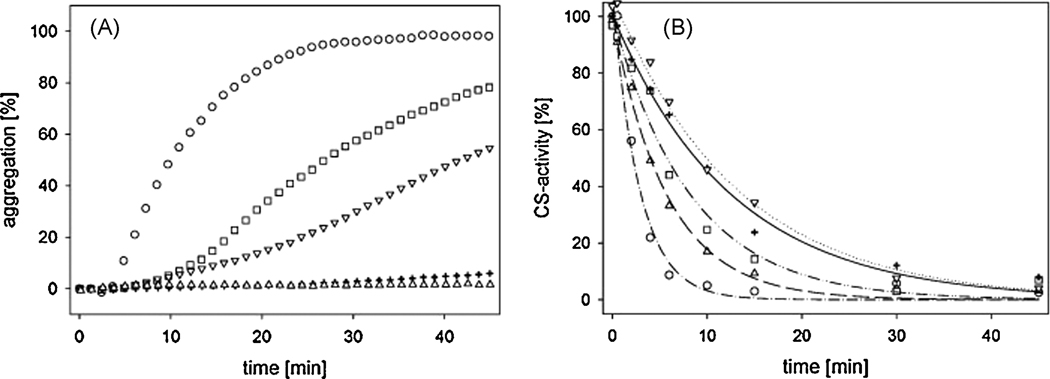
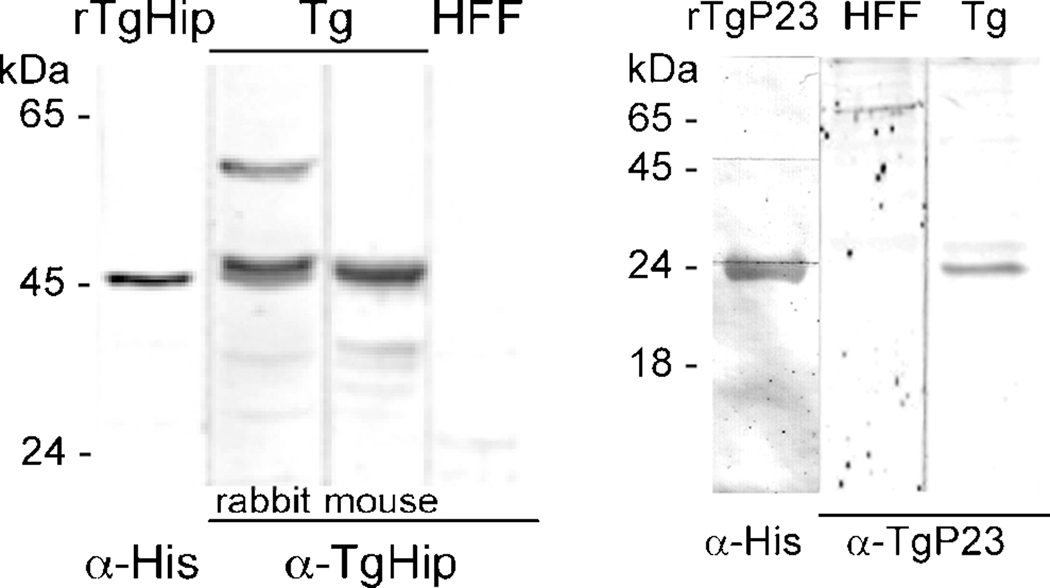
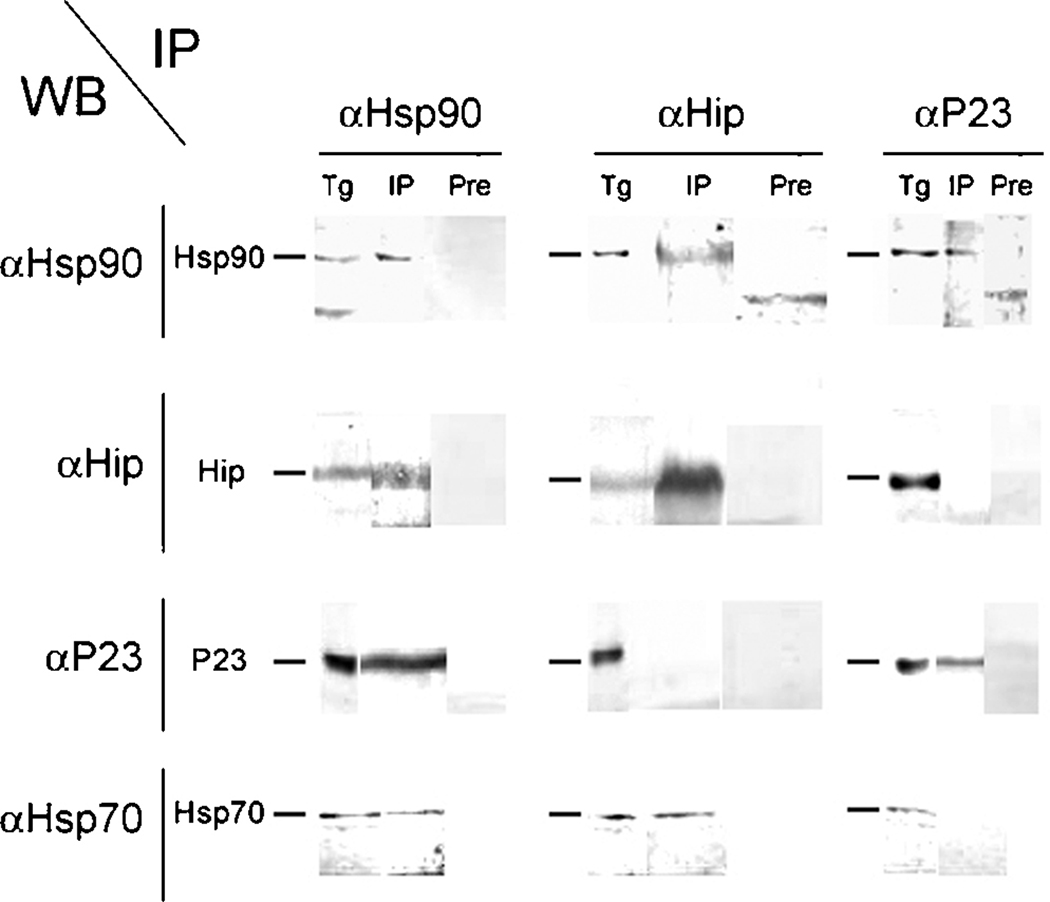
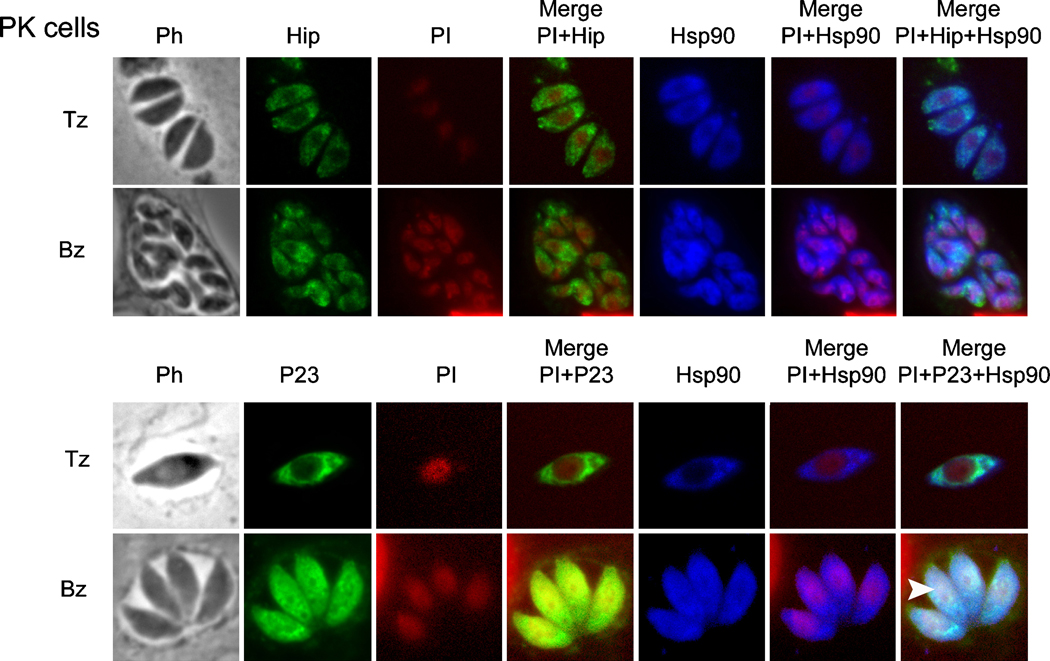
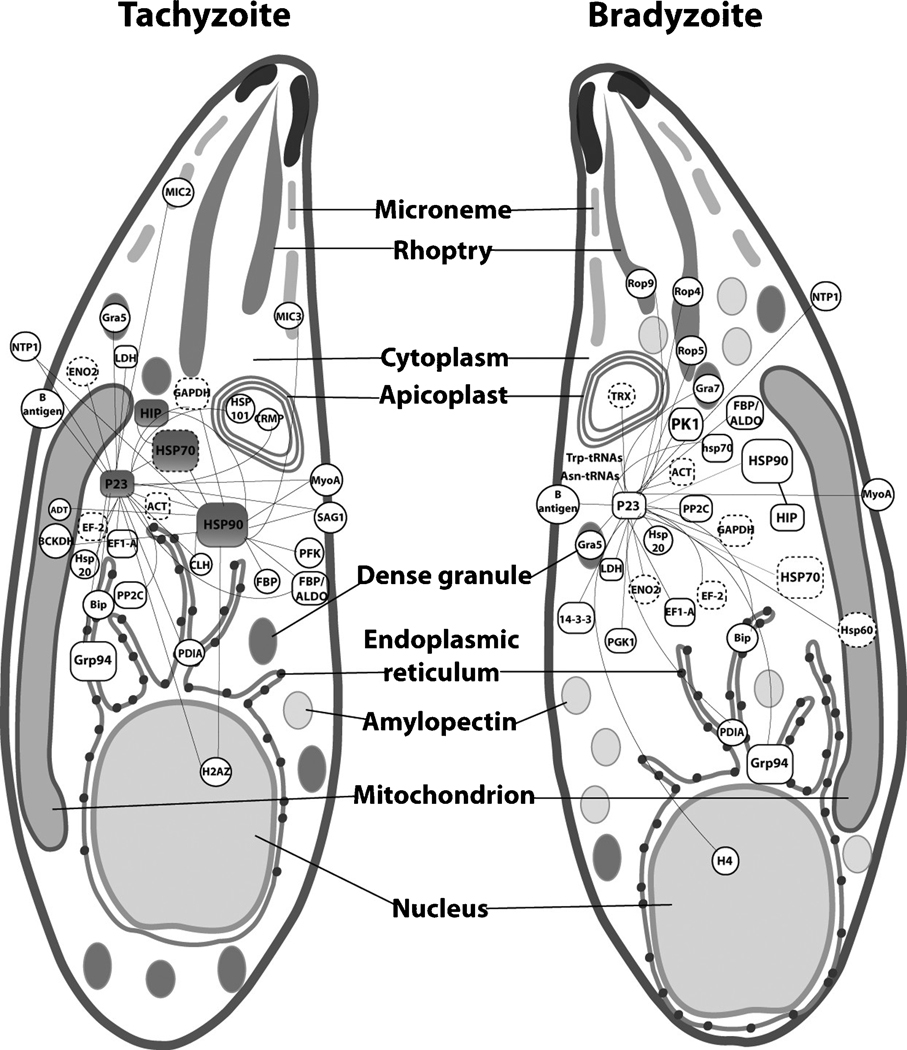
Toxoplasma gondii is among the most successful parasites, with nearly half of the human population chronically infected. Recently a link between the T. gondii Hsp90 chaperone machinery and parasite development was observed. Here, the T. gondii Hsp90 co-chaperones p23 and Hip were identified mining the Toxoplasma- database (www.toxodb.org). Their identity was confirmed by domain structure and blast analysis. Additionally, analysis of the secondary structure and studies on the chaperone function of the purified protein verified the p23 identity. Studies of co-immunoprecipitation (co-IP) identified two different types of complexes, one comprising at least Hip-Hsp70-Hsp90 and another containing at least p23-Hsp90. Indirect immunofluorescence assays showed that Hip is localized in the cytoplasm in tachyzoites and as well in bradyzoites. For p23 in contrast, a solely cytoplasmic localization was only observed in the tachyzoite stage whereas nuclear and cytosolic distribution and co-localization with Hsp90 was observed in bradyzoites. These results indicate that the T. gondii Hsp90-heterocomplex cycle is similar to the one proposed for higher eukaryotes, further highlighting the implication of the Hsp90/p23 in parasite development. Furthermore, co-IP experiments of tachyzoite/bradyzoite lysates with anti-p23 antiserum and identification of the complexed proteins together with the use of the curated interaction data available from different source (orthologs and Plasmodium databases) allowed us to construct an interaction network (interactome) covering the dynamics of the Hsp90 chaperone machinery.
Figures
References
-
- Ferreira da Silva Mda F, Barbosa HS, Gross U, Luder CG. Stress-related and spontaneous stage differentiation of Toxoplasma gondii. Mol Biosyst. 2008;4:824–834. - PubMed
-
- Echeverria PC, Matrajt M, Harb OS, et al. Toxoplasma gondii Hsp90 is a potential drug target whose expression and subcellular localization are developmentally regulated. J Mol Biol. 2005;350:723–734. - PubMed
-
- Ahn HJ, Kim S, Nam HW. Molecular cloning of the 82-kDa heat shock protein (HSP90) of Toxoplasma gondii associated with the entry into and growth in host cells. Biochem Biophys Res Commun. 2003;311:654–659. - PubMed
-
- Jakob U, Lilie H, Meyer I, Buchner J. Transient interaction of hsp90 with early unfolding intermediates of citrate synthase - implications for heat shock in vivo. J Biol Chem. 1995;270:7288–7294. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
