

Expression of microRNAs and its regulation in plants
- PMID: 20403450
- PMCID: PMC2939293
- DOI: 10.1016/j.semcdb.2010.03.012
Expression of microRNAs and its regulation in plants
Abstract
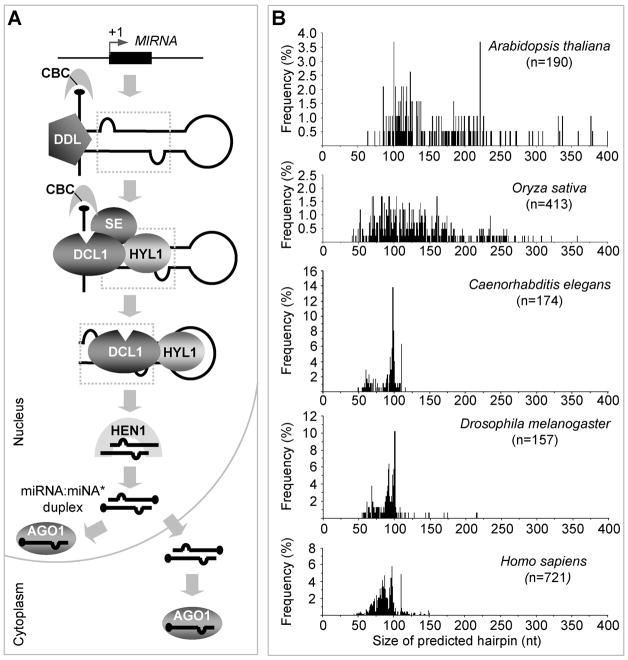
MicroRNAs (miRNAs) have emerged as an essential regulatory component in plants. Many of the known miRNAs are evolutionarily conserved across diverse plant species and function in the regulatory control of fundamentally important biological processes such as developmental timing, patterning, and response to environmental changes. Expression of miRNAs in plants involves transcription from MIRNA loci by RNA polymerase II (pol II), multi-step processing of the primary transcripts by the DICER-LIKE1 (DCL1) complex, and formation of effector complexes consisting of mature miRNAs and ARGONAUTE (AGO) family proteins. In this short review, we present the most recent advances in our understanding of the molecular machinery as well as the regulatory mechanisms involved in the expression of miRNAs in plants.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures

Similar articles
-
NOT2 proteins promote polymerase II-dependent transcription and interact with multiple MicroRNA biogenesis factors in Arabidopsis.Plant Cell. 2013 Feb;25(2):715-27. doi: 10.1105/tpc.112.105882. Epub 2013 Feb 19. Plant Cell. 2013. PMID: 23424246 Free PMC article.
-
Recent advances in the regulation of plant miRNA biogenesis.RNA Biol. 2021 Dec;18(12):2087-2096. doi: 10.1080/15476286.2021.1899491. Epub 2021 Mar 17. RNA Biol. 2021. PMID: 33666136 Free PMC article. Review.
-
Biogenesis, turnover, and mode of action of plant microRNAs.Plant Cell. 2013 Jul;25(7):2383-99. doi: 10.1105/tpc.113.113159. Epub 2013 Jul 23. Plant Cell. 2013. PMID: 23881412 Free PMC article. Review.
-
Bidirectional processing of pri-miRNAs with branched terminal loops by Arabidopsis Dicer-like1.Nat Struct Mol Biol. 2013 Sep;20(9):1106-15. doi: 10.1038/nsmb.2646. Epub 2013 Aug 11. Nat Struct Mol Biol. 2013. PMID: 23934148 Free PMC article.
-
New insights into pri-miRNA processing and accumulation in plants.Wiley Interdiscip Rev RNA. 2015 Sep-Oct;6(5):533-45. doi: 10.1002/wrna.1292. Epub 2015 Jun 29. Wiley Interdiscip Rev RNA. 2015. PMID: 26119101 Review.
Cited by
-
In Arabidopsis thaliana, RNA-Induced Silencing Complex-Loading of MicroRNAs Plays a Minor Regulatory Role During Photomorphogenesis Except for miR163.Front Plant Sci. 2022 Jul 13;13:854869. doi: 10.3389/fpls.2022.854869. eCollection 2022. Front Plant Sci. 2022. PMID: 35909792 Free PMC article.
-
NOT2 proteins promote polymerase II-dependent transcription and interact with multiple MicroRNA biogenesis factors in Arabidopsis.Plant Cell. 2013 Feb;25(2):715-27. doi: 10.1105/tpc.112.105882. Epub 2013 Feb 19. Plant Cell. 2013. PMID: 23424246 Free PMC article.
-
Identification and characterisation of microRNAs and their target genes in phosphate-starved Nicotiana benthamiana by small RNA deep sequencing and 5'RACE analysis.BMC Genomics. 2018 Dec 17;19(1):940. doi: 10.1186/s12864-018-5258-9. BMC Genomics. 2018. PMID: 30558535 Free PMC article.
-
Hyponastic Leaves 1 protects pri-miRNAs from nuclear exosome attack.Proc Natl Acad Sci U S A. 2020 Jul 21;117(29):17429-17437. doi: 10.1073/pnas.2007203117. Epub 2020 Jul 7. Proc Natl Acad Sci U S A. 2020. PMID: 32636270 Free PMC article.
-
Miniature Inverted-Repeat Transposable Elements: Small DNA Transposons That Have Contributed to Plant MICRORNA Gene Evolution.Plants (Basel). 2023 Mar 1;12(5):1101. doi: 10.3390/plants12051101. Plants (Basel). 2023. PMID: 36903960 Free PMC article. Review.
References
-
- Chapman EJ, Carrington JC. Specialization and evolution of endogenous small RNA pathways. Nat Rev Genet. 2007;8:884–96. - PubMed
-
- Jones-Rhoades MW, Bartel DP, Bartel B. MicroRNAs and their Regulatory Roles in Plants. Annu Rev Plant Biol. 2006;57:19–53. - PubMed
-
- Voinnet O. Origin, biogenesis, and activity of plant microRNAs. Cell. 2009;136:669–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
