

KLF3 regulates muscle-specific gene expression and synergizes with serum response factor on KLF binding sites
- PMID: 20404088
- PMCID: PMC2897560
- DOI: 10.1128/MCB.00302-10
KLF3 regulates muscle-specific gene expression and synergizes with serum response factor on KLF binding sites
Abstract
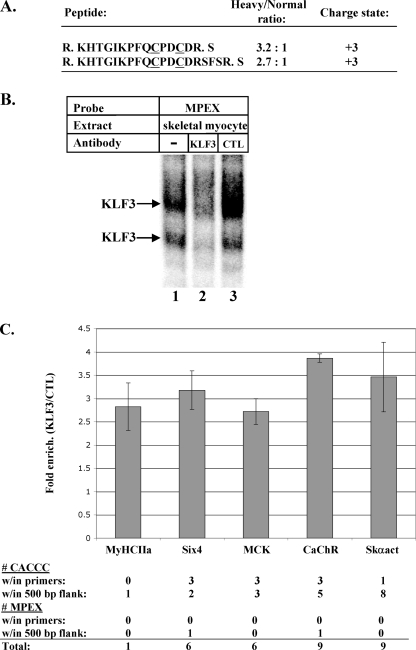
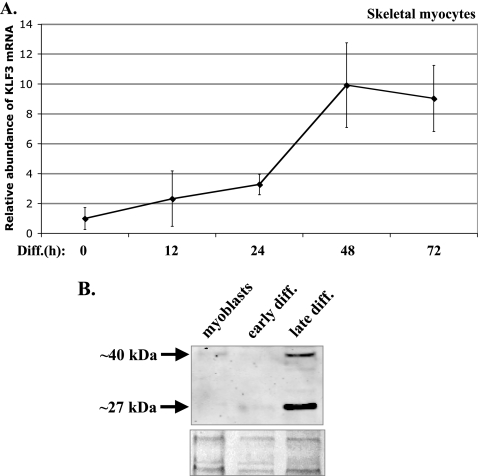
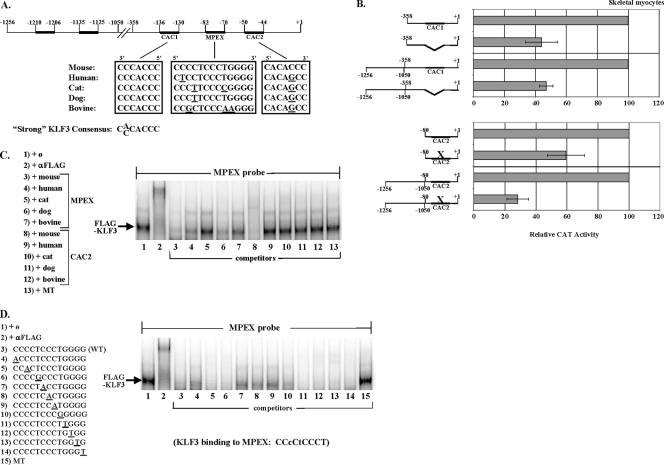
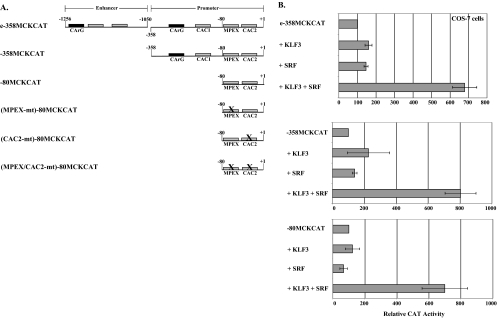
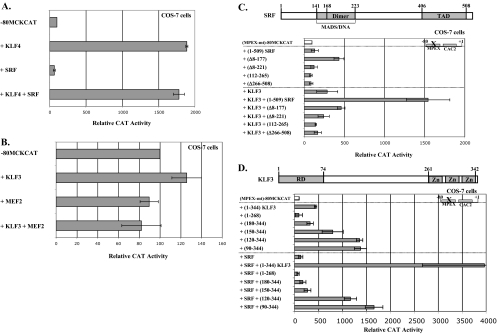
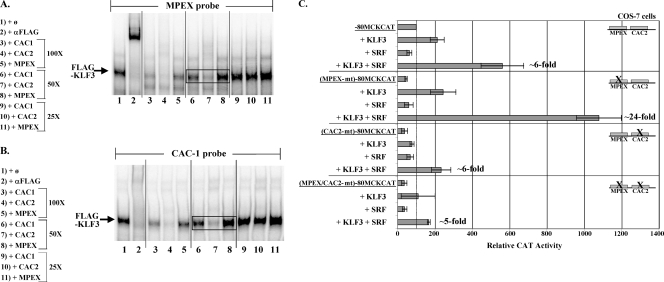
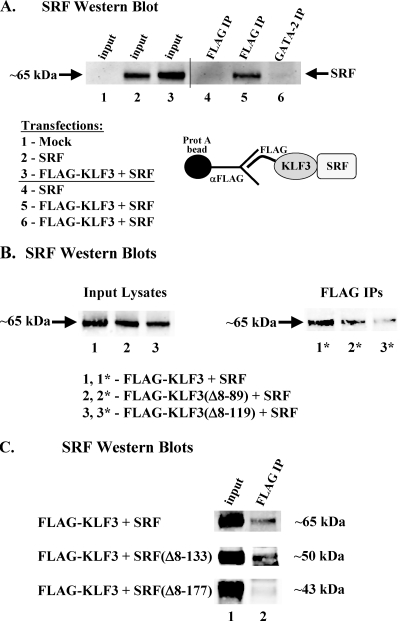
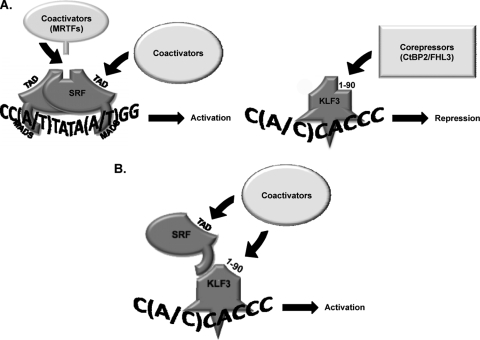
This study identifies KLF3 as a transcriptional regulator of muscle genes and reveals a novel synergistic interaction between KLF3 and serum response factor (SRF). Using quantitative proteomics, KLF3 was identified as one of several candidate factors that recognize the MPEX control element in the Muscle creatine kinase (MCK) promoter. Chromatin immunoprecipitation analysis indicated that KLF3 is enriched at many muscle gene promoters (MCK, Myosin heavy chain IIa, Six4, Calcium channel receptor alpha-1, and Skeletal alpha-actin), and two KLF3 isoforms are upregulated during muscle differentiation. KLF3 and SRF physically associate and synergize in transactivating the MCK promoter independently of SRF binding to CArG motifs. The zinc finger and repression domains of KLF3 plus the MADS box and transcription activation domain of SRF are implicated in this synergy. Our results provide the first evidence of a role for KLF3 in muscle gene regulation and reveal an alternate mechanism for transcriptional regulation by SRF via its recruitment to KLF binding sites. Since both factors are expressed in all muscle lineages, SRF may regulate many striated- and smooth-muscle genes that lack known SRF control elements, thus further expanding the breadth of the emerging CArGome.
Figures
Similar articles
-
Quantitative proteomic identification of MAZ as a transcriptional regulator of muscle-specific genes in skeletal and cardiac myocytes.Mol Cell Biol. 2008 Oct;28(20):6521-35. doi: 10.1128/MCB.00306-08. Epub 2008 Aug 18. Mol Cell Biol. 2008. PMID: 18710939 Free PMC article.
-
Leukocyte β7 integrin targeted by Krüppel-like factors.J Immunol. 2014 Aug 15;193(4):1737-46. doi: 10.4049/jimmunol.1302613. Epub 2014 Jul 11. J Immunol. 2014. PMID: 25015818
-
Serum response factor MADS box serine-162 phosphorylation switches proliferation and myogenic gene programs.Proc Natl Acad Sci U S A. 2006 Mar 21;103(12):4516-21. doi: 10.1073/pnas.0505338103. Epub 2006 Mar 13. Proc Natl Acad Sci U S A. 2006. PMID: 16537394 Free PMC article.
-
The mammalian zinc finger transcription factor Krüppel-like factor 3 (KLF3/BKLF).IUBMB Life. 2011 Feb;63(2):86-93. doi: 10.1002/iub.422. Epub 2011 Feb 24. IUBMB Life. 2011. PMID: 21360637 Review.
-
Serum response factor: toggling between disparate programs of gene expression.J Mol Cell Cardiol. 2003 Jun;35(6):577-93. doi: 10.1016/s0022-2828(03)00110-x. J Mol Cell Cardiol. 2003. PMID: 12788374 Review.
Cited by
-
miR-21-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting KLF3 in Chicken.Genes (Basel). 2021 May 26;12(6):814. doi: 10.3390/genes12060814. Genes (Basel). 2021. PMID: 34073601 Free PMC article.
-
Identification and expression pattern analysis of miRNAs in pectoral muscle during pigeon (Columba livia) development.PeerJ. 2021 Jun 23;9:e11438. doi: 10.7717/peerj.11438. eCollection 2021. PeerJ. 2021. PMID: 34221709 Free PMC article.
-
Differential regulation of the α-globin locus by Krüppel-like Factor 3 in erythroid and non-erythroid cells.BMC Mol Biol. 2014 May 16;15:8. doi: 10.1186/1471-2199-15-8. BMC Mol Biol. 2014. PMID: 24885809 Free PMC article.
-
Bone mesenchymal stem cells (BMSCs)-derived exosomal microRNA-21-5p regulates Kruppel-like factor 3 (KLF3) to promote osteoblast proliferation in vitro.Bioengineered. 2022 May;13(5):11933-11944. doi: 10.1080/21655979.2022.2067286. Bioengineered. 2022. PMID: 35549815 Free PMC article.
-
Muscle-relevant genes marked by stable H3K4me2/3 profiles and enriched MyoD binding during myogenic differentiation.PLoS One. 2017 Jun 13;12(6):e0179464. doi: 10.1371/journal.pone.0179464. eCollection 2017. PLoS One. 2017. PMID: 28609469 Free PMC article.
References
-
- Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 72:248-254. - PubMed
-
- Camoretti-Mercado, B., D. J. Fernandes, S. Dewundara, J. Churchill, L. Ma, P. C. Kogut, J. F. McConville, M. S. Parmacek, and J. Solway. 2006. Inhibition of transforming growth factor beta-enhanced serum response factor-dependent transcription by SMAD7. J. Biol. Chem. 281:20383-20392. - PubMed
-
- Cen, B., A. Selvaraj, and R. Prywes. 2004. Myocardin/MKL family of SRF coactivators: key regulators of immediate early and muscle specific gene expression. J. Cell. Biochem. 93:74-82. - PubMed
-
- Chang, D. F., N. S. Belaguli, D. Iyer, W. B. Roberts, S. P. Wu, X. R. Dong, J. G. Marx, M. S. Moore, M. C. Beckerle, M. W. Majesky, and R. J. Schwartz. 2003. Cysteine-rich LIM-only proteins CRP1 and CRP2 are potent smooth muscle differentiation cofactors. Dev. Cell 4:107-118. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous