

Transcriptional activity affects the H3K4me3 level and distribution in the coding region
- PMID: 20404096
- PMCID: PMC2876678
- DOI: 10.1128/MCB.01478-09
Transcriptional activity affects the H3K4me3 level and distribution in the coding region
Abstract
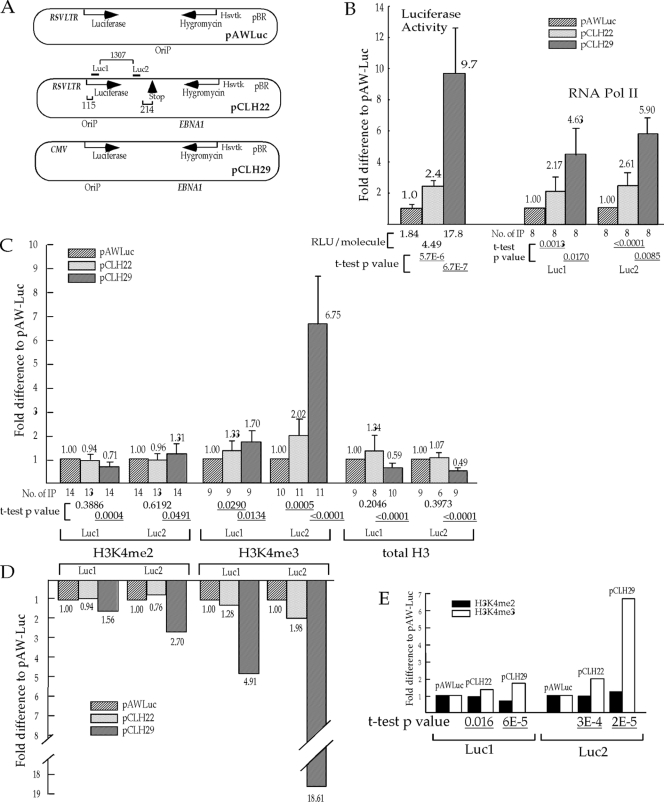
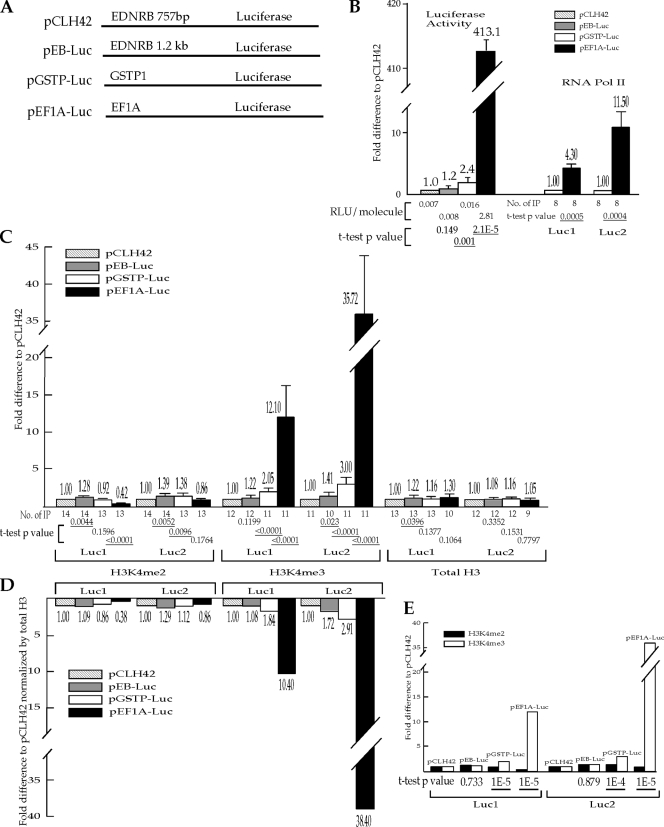
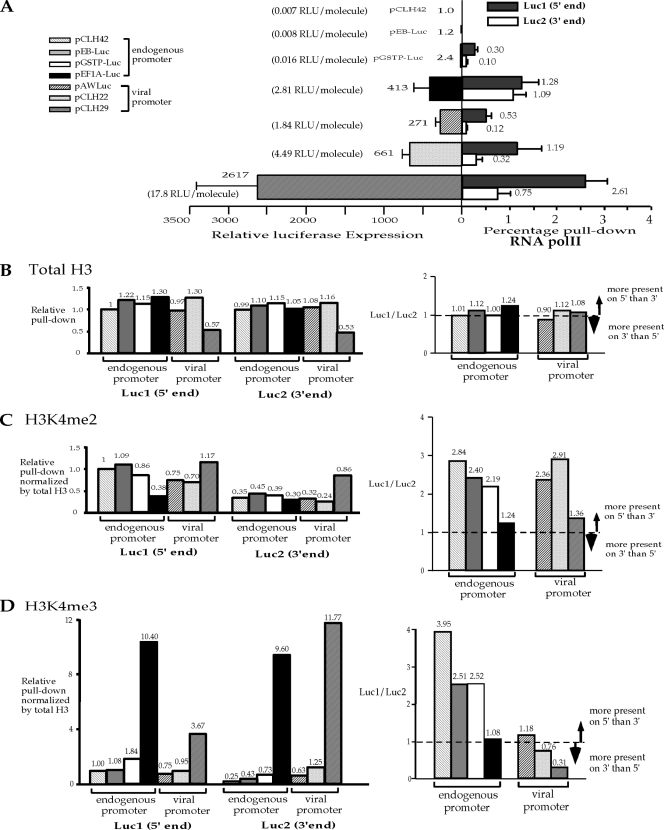
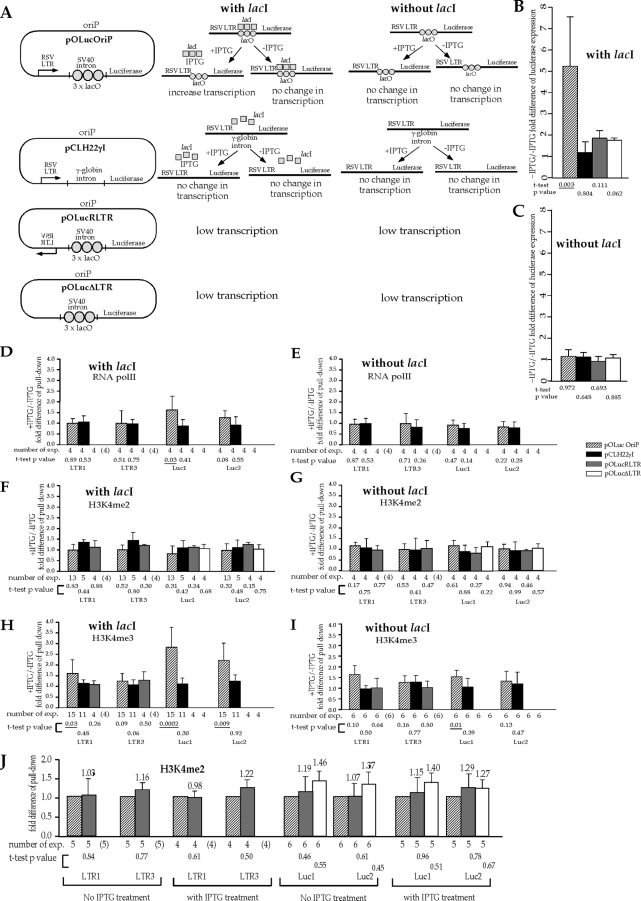
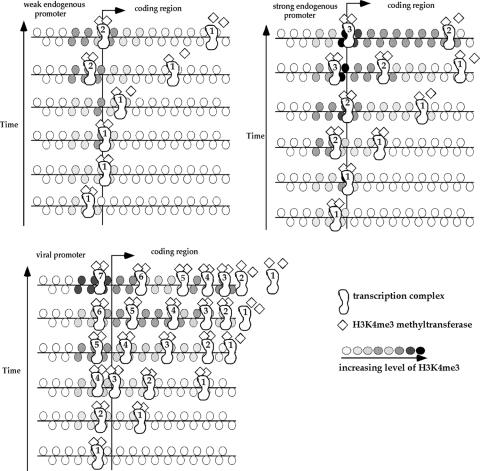
Histone lysine methylation and CpG DNA methylation contribute to transcriptional regulation. We have shown previously that dimethylated and trimethylated forms of histone H3 at lysine 4 (H3K4me2 and H3K4me3) are primarily depleted from CpG-methylated DNA regions by using patch-methylated stable episomes (minichromosomes) in human cells. This effect on H3K4me2 is clearly not linked to the transcriptional activity in the methylated DNA region; however, transcriptional activity may play a role in the presence of H3K4me3. Here, we present clear evidence of the impact of transcriptional activity on the overall level of H3K4me3 in the coding region and the lack of impact on H3K4me2. Our data also demonstrate the influence of transcriptional activity on the distribution of H3K4me3 and H3K4me2, but not that of total H3, in the 5' end of the coding region relative to the 3' end. The nature of the promoter (viral or endogenous) affects H3K4me3 much more than it affects H3K4me2, suggesting a potential fundamental difference in the recruitment of methyltransferase for H3K4 trimethylation.
Figures
References
-
- Allis, C. D., S. L. Berger, J. Cote, S. Dent., T. Jenuwien, T. Kouzarides, L. Pillus, D. Reinberg, Y. Shi, R. Shiekhattar, A. Shilatifard, J. Workman, and Y. Zhang. 2007. New nomenclature for chromatin-modifying enzymes. Cell 131:633-636. - PubMed
-
- Barski, A., S. Cuddapah, K. Cui, T. Y. Roh, D. E. Schones, Z. Wang, G. Wei, I. Chepelev, and K. Zhao. 2007. High-resolution profiling of histone methylations in the human genome. Cell 129:823-837. - PubMed
-
- Bernstein, B. E., M. Kamal, K. Lindblad-Toh, S. Bekiranov, D. K. Bailey, D. J. Huebert, S. McMahon, E. K. Karlsson, E. J. Kulbokas III, T. R. Gingeras, S. L. Schreiber, and E. S. Lander. 2005. Genomic maps and comparative analysis of histone modifications in human and mouse. Cell 120:169-181. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases