

Regulation of exosome secretion by Rab35 and its GTPase-activating proteins TBC1D10A-C
- PMID: 20404108
- PMCID: PMC2856897
- DOI: 10.1083/jcb.200911018
Regulation of exosome secretion by Rab35 and its GTPase-activating proteins TBC1D10A-C
Abstract
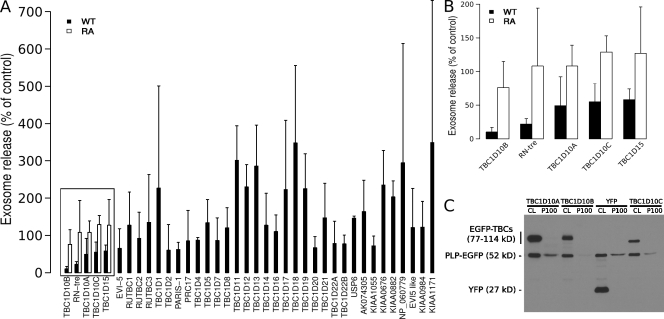
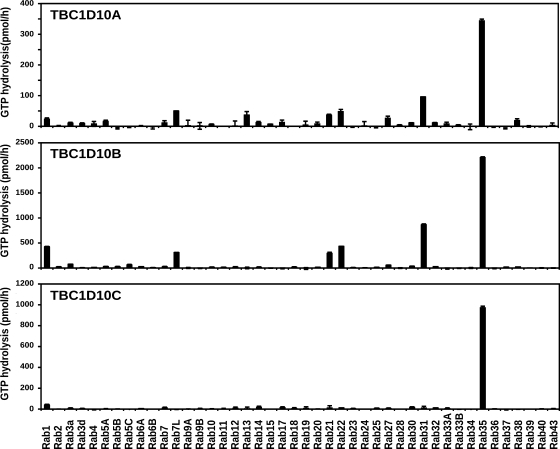
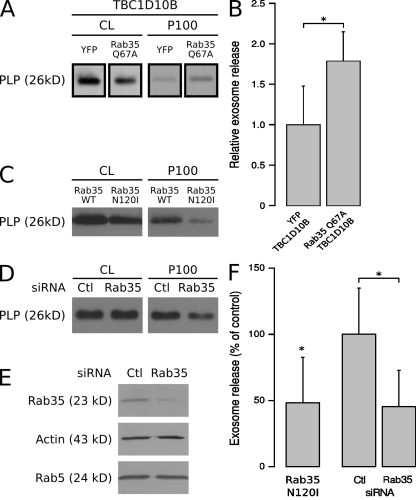
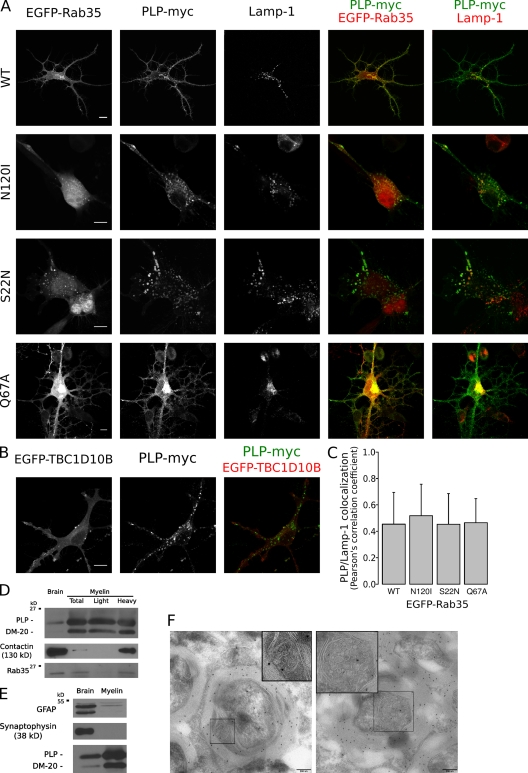
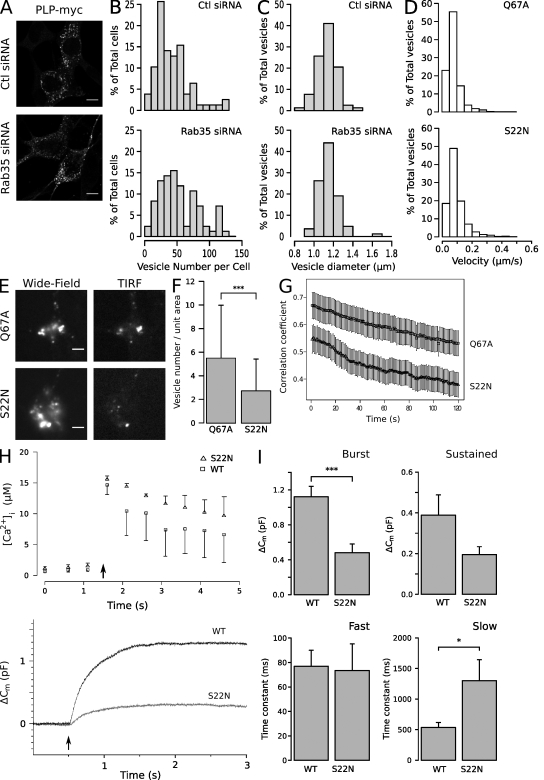
Oligodendrocytes secrete vesicles into the extracellular space, where they might play a role in neuron-glia communication. These exosomes are small vesicles with a diameter of 50-100 nm that are formed within multivesicular bodies and are released after fusion with the plasma membrane. The intracellular pathways that generate exosomes are poorly defined. Because Rab family guanosine triphosphatases (GTPases) together with their regulators are important membrane trafficking organizers, we investigated which Rab GTPase-activating proteins interfere with exosome release. We find that TBC1D10A-C regulate exosome secretion in a catalytic activity-dependent manner. We show that Rab35 is the target of TBC1D10A-C and that the inhibition of Rab35 function leads to intracellular accumulation of endosomal vesicles and impairs exosome secretion. Rab35 localizes to the surface of oligodendroglia in a GTP-dependent manner, where it increases the density of vesicles, suggesting a function in docking or tethering. These findings provide a basis for understanding the biogenesis and function of exosomes in the central nervous system.
Figures
Similar articles
-
Rab35 protein regulates evoked exocytosis of endothelial Weibel-Palade bodies.J Biol Chem. 2017 Jul 14;292(28):11631-11640. doi: 10.1074/jbc.M116.773333. Epub 2017 May 31. J Biol Chem. 2017. PMID: 28566286 Free PMC article.
-
Rab27A regulates exosome secretion from lung adenocarcinoma cells A549: involvement of EPI64.APMIS. 2014 Nov;122(11):1080-7. doi: 10.1111/apm.12261. Epub 2014 Mar 28. APMIS. 2014. PMID: 24673604
-
Long non-coding RNA HOTAIR promotes exosome secretion by regulating RAB35 and SNAP23 in hepatocellular carcinoma.Mol Cancer. 2019 Apr 3;18(1):78. doi: 10.1186/s12943-019-0990-6. Mol Cancer. 2019. PMID: 30943982 Free PMC article.
-
Rab35: GEFs, GAPs and effectors.Traffic. 2013 Nov;14(11):1109-17. doi: 10.1111/tra.12096. Epub 2013 Aug 21. Traffic. 2013. PMID: 23905989 Review.
-
New insights into the function of Rab GTPases in the context of exosomal secretion.Small GTPases. 2018 Mar 4;9(1-2):95-106. doi: 10.1080/21541248.2016.1264352. Epub 2017 Jan 31. Small GTPases. 2018. PMID: 28135905 Free PMC article. Review.
Cited by
-
Focus on the morphogenesis, fate and the role in tumor progression of multivesicular bodies.Cell Commun Signal. 2020 Aug 8;18(1):122. doi: 10.1186/s12964-020-00619-5. Cell Commun Signal. 2020. PMID: 32771015 Free PMC article. Review.
-
Exosome and its roles in cardiovascular diseases.Heart Fail Rev. 2015 May;20(3):337-48. doi: 10.1007/s10741-014-9469-0. Heart Fail Rev. 2015. PMID: 25549884 Review.
-
Microglia: Agents of the CNS Pro-Inflammatory Response.Cells. 2020 Jul 17;9(7):1717. doi: 10.3390/cells9071717. Cells. 2020. PMID: 32709045 Free PMC article. Review.
-
The Role of Exosomes in Stemness and Neurodegenerative Diseases-Chemoresistant-Cancer Therapeutics and Phytochemicals.Int J Mol Sci. 2020 Sep 17;21(18):6818. doi: 10.3390/ijms21186818. Int J Mol Sci. 2020. PMID: 32957534 Free PMC article. Review.
-
Exosomes in cancer development, metastasis, and drug resistance: a comprehensive review.Cancer Metastasis Rev. 2013 Dec;32(3-4):623-42. doi: 10.1007/s10555-013-9441-9. Cancer Metastasis Rev. 2013. PMID: 23709120 Free PMC article. Review.
References
-
- Aoki N., Jin-no S., Nakagawa Y., Asai N., Arakawa E., Tamura N., Tamura T., Matsuda T. 2007. Identification and characterization of microvesicles secreted by 3T3-L1 adipocytes: redox- and hormone-dependent induction of milk fat globule-epidermal growth factor 8-associated microvesicles. Endocrinology. 148:3850–3862 10.1210/en.2006-1479 - DOI - PubMed
-
- Buschow S.I., Nolte-’t Hoen E.N., van Niel G., Pols M.S., ten Broeke T., Lauwen M., Ossendorp F., Melief C.J., Raposo G., Wubbolts R., et al. 2009. MHC II in dendritic cells is targeted to lysosomes or T cell-induced exosomes via distinct multivesicular body pathways. Traffic. 10:1528–1542 10.1111/j.1600-0854.2009.00963.x - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
