

Drosophila TRPA1 channel mediates chemical avoidance in gustatory receptor neurons
- PMID: 20404155
- PMCID: PMC2889570
- DOI: 10.1073/pnas.1001425107
Drosophila TRPA1 channel mediates chemical avoidance in gustatory receptor neurons
Abstract
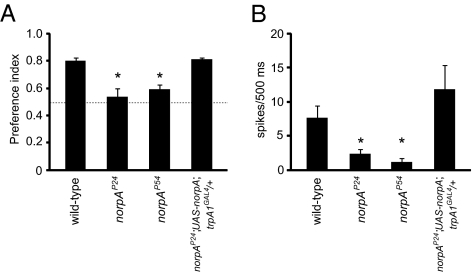
Mammalian sweet, bitter, and umami taste is mediated by a single transduction pathway that includes a phospholipase C (PLC)beta and one cation channel, TRPM5. However, in insects such as the fruit fly, Drosophila melanogaster, it is unclear whether different tastants, such as bitter compounds, are sensed in gustatory receptor neurons (GRNs) through one or multiple ion channels, as the cation channels required in insect GRNs are unknown. Here, we set out to explore additional sensory roles for the Drosophila TRPA1 channel, which was known to function in thermosensation. We found that TRPA1 was expressed in GRNs that respond to aversive compounds. Elimination of TRPA1 had no impact on the responses to nearly all bitter compounds tested, including caffeine, quinine, and strychnine. Rather, we found that TRPA1 was required in a subset of avoidance GRNs for the behavioral and electrophysiological responses to aristolochic acid. TRPA1 did not appear to be activated or inhibited directly by aristolochic acid. We found that elimination of the same PLC that leads to activation of TRPA1 in thermosensory neurons was also required in the TRPA1-expressing GRNs for avoiding aristolochic acid. Given that mammalian TRPA1 is required for responding to noxious chemicals, many of which cause pain and injury, our analysis underscores the evolutionarily conserved role for TRPA1 channels in chemical avoidance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
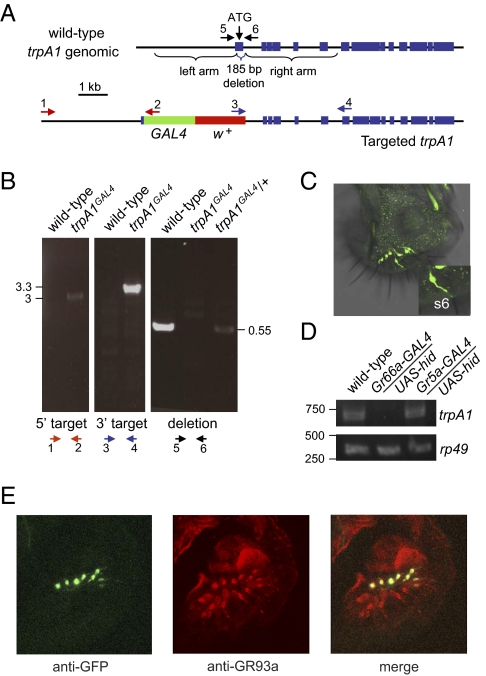
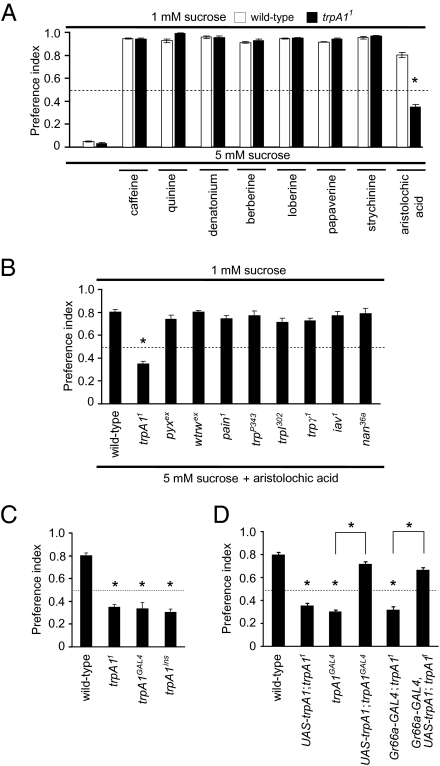
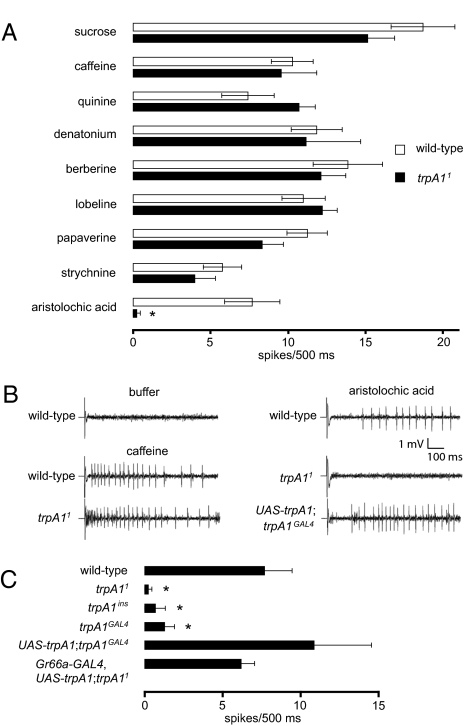
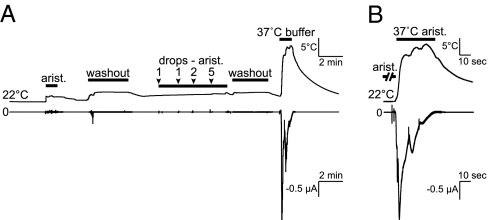
Similar articles
-
Modulation of TRPA1 thermal sensitivity enables sensory discrimination in Drosophila.Nature. 2011 Dec 4;481(7379):76-80. doi: 10.1038/nature10715. Nature. 2011. PMID: 22139422 Free PMC article.
-
H2O2-Sensitive Isoforms of Drosophila melanogaster TRPA1 Act in Bitter-Sensing Gustatory Neurons to Promote Avoidance of UV During Egg-Laying.Genetics. 2017 Feb;205(2):749-759. doi: 10.1534/genetics.116.195172. Epub 2016 Dec 7. Genetics. 2017. PMID: 27932542 Free PMC article.
-
A gustatory receptor paralogue controls rapid warmth avoidance in Drosophila.Nature. 2013 Aug 29;500(7464):580-4. doi: 10.1038/nature12390. Epub 2013 Aug 7. Nature. 2013. PMID: 23925112 Free PMC article.
-
Temperature sensation in Drosophila.Curr Opin Neurobiol. 2015 Oct;34:8-13. doi: 10.1016/j.conb.2015.01.002. Epub 2015 Jan 21. Curr Opin Neurobiol. 2015. PMID: 25616212 Free PMC article. Review.
-
Drosophila sensory receptors-a set of molecular Swiss Army Knives.Genetics. 2021 Mar 3;217(1):1-34. doi: 10.1093/genetics/iyaa011. Genetics. 2021. PMID: 33683373 Free PMC article. Review.
Cited by
-
TRPA1 mediates sensation of the rate of temperature change in Drosophila larvae.Nat Neurosci. 2017 Jan;20(1):34-41. doi: 10.1038/nn.4416. Epub 2016 Oct 17. Nat Neurosci. 2017. PMID: 27749829 Free PMC article.
-
The transient receptor potential channel TRPA1: from gene to pathophysiology.Pflugers Arch. 2012 Nov;464(5):425-58. doi: 10.1007/s00424-012-1158-z. Epub 2012 Sep 22. Pflugers Arch. 2012. PMID: 23001121 Review.
-
TRP Channels as Sensors of Bacterial Endotoxins.Toxins (Basel). 2018 Aug 11;10(8):326. doi: 10.3390/toxins10080326. Toxins (Basel). 2018. PMID: 30103489 Free PMC article. Review.
-
Drosophila TRPA1 channel is required to avoid the naturally occurring insect repellent citronellal.Curr Biol. 2010 Sep 28;20(18):1672-8. doi: 10.1016/j.cub.2010.08.016. Curr Biol. 2010. PMID: 20797863 Free PMC article.
-
Drosophila gustatory preference behaviors require the atypical soluble guanylyl cyclases.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2011 Jul;197(7):717-27. doi: 10.1007/s00359-011-0634-9. Epub 2011 Feb 25. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2011. PMID: 21350862
References
-
- Story GM, et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell. 2003;112:819–829. - PubMed
-
- Viswanath V, et al. Opposite thermosensor in fruitfly and mouse. Nature. 2003;423:822–823. - PubMed
-
- Kindt KS, et al. Caenorhabditis elegans TRPA-1 functions in mechanosensation. Nat Neurosci. 2007;10:568–577. - PubMed
-
- Bandell M, et al. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron. 2004;41:849–857. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
