

SKI promotes Smad3 linker phosphorylations associated with the tumor-promoting trait of TGFbeta
- PMID: 20404506
- PMCID: PMC3235508
- DOI: 10.4161/cc.9.9.11292
SKI promotes Smad3 linker phosphorylations associated with the tumor-promoting trait of TGFbeta
Abstract
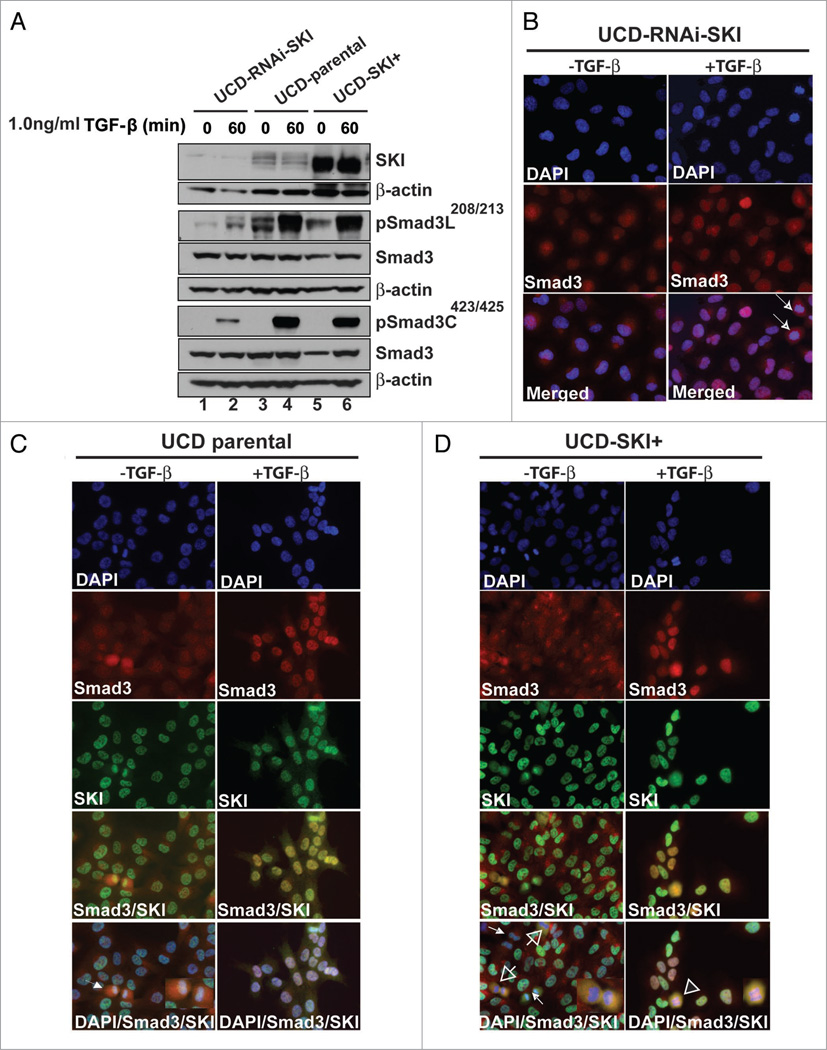
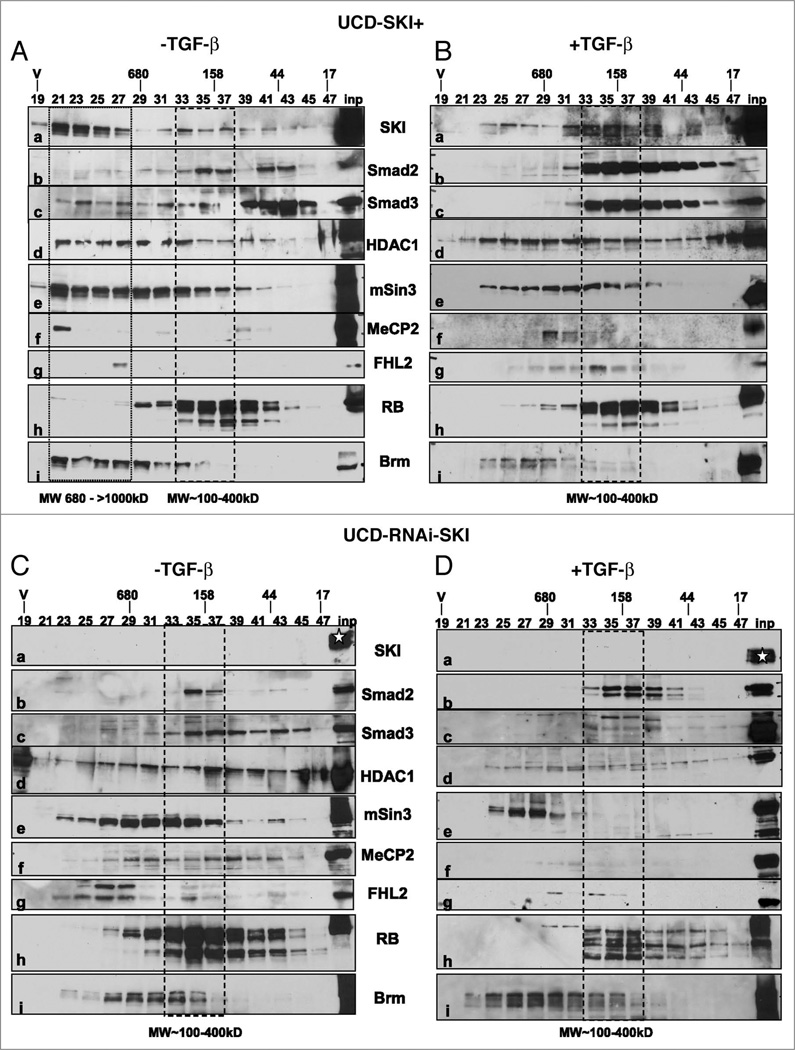
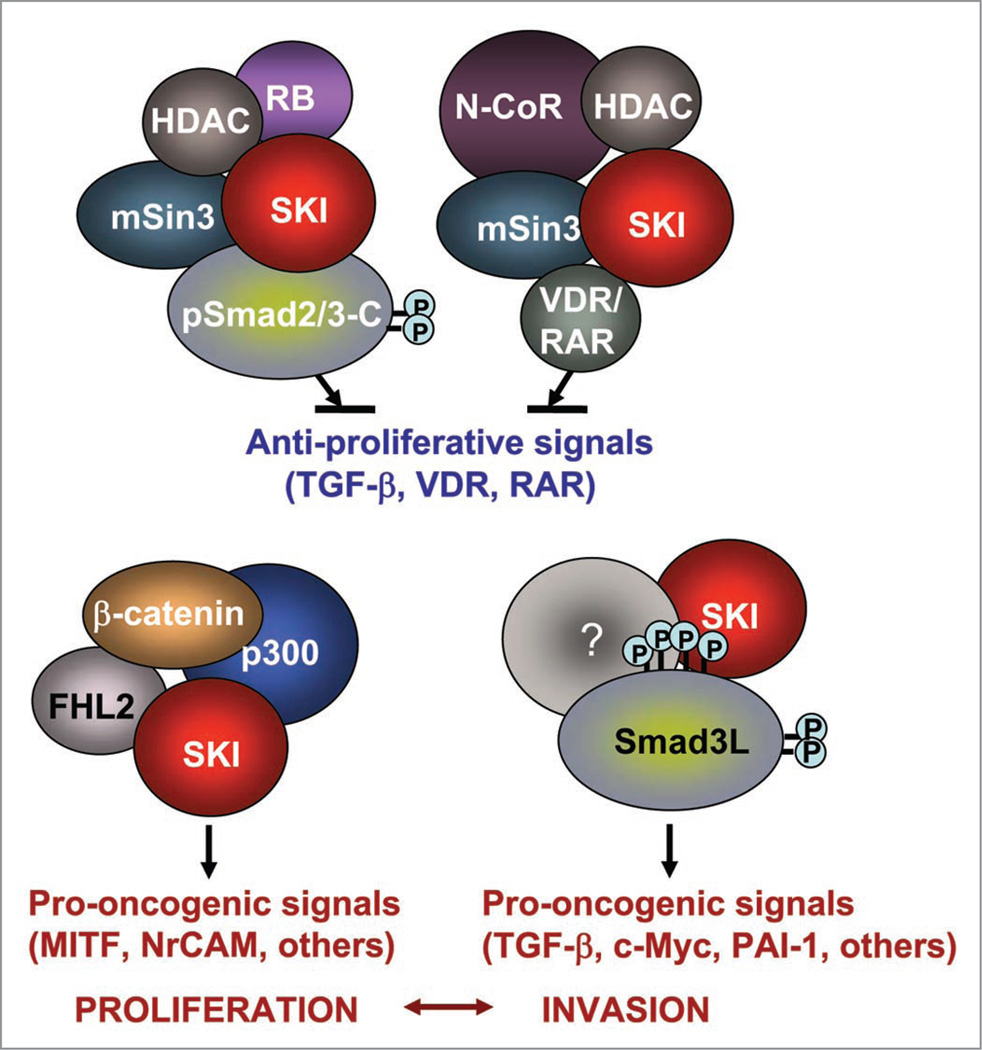
The transcriptional co-regulator SKI is a potent inhibitor of TGFbeta-growth inhibitory signals. SKI binds to receptor-activated Smads in the nucleus, forming repressor complexes containing HDACs, mSin3, NCoR, and other protein partners. Alternatively, SKI binds to activated Smads in the cytoplasm, preventing their nuclear translocation. SKI is necessary for anchorage-independent growth of melanoma cells in vitro, and most important, for human melanoma xenograft growth in vivo. We recently identified a novel role of SKI in TGFbeta signaling. SKI promotes the switch of Smad3 from repressor of proliferation to activator of oncogenesis by facilitating phosphorylations in the linker domain. High levels of endogenous SKI are required by the tumor promoting trait of TGFbeta to induce expression of the plasminogen-activator inhibitor-1 (PAI-1), sustained expression of C-Myc and for aborting upregulation of p21(Waf-1). Here we discuss how SKI diversifies and amplifies its functions by associating with multiple protein partners and by promoting Smad3 linker phosphorylation(s) in response to TGFbeta signaling in melanoma cells.
Figures
Similar articles
-
SKI knockdown inhibits human melanoma tumor growth in vivo.Pigment Cell Melanoma Res. 2009 Dec;22(6):761-72. doi: 10.1111/j.1755-148X.2009.00603.x. Pigment Cell Melanoma Res. 2009. PMID: 19845874 Free PMC article.
-
Repression of TGF-beta signaling by the oncogenic protein SKI in human melanomas: consequences for proliferation, survival, and metastasis.Oncogene. 2003 May 19;22(20):3123-9. doi: 10.1038/sj.onc.1206452. Oncogene. 2003. PMID: 12793438 Review.
-
Efficient TGF-β/SMAD signaling in human melanoma cells associated with high c-SKI/SnoN expression.Mol Cancer. 2011 Jan 6;10(1):2. doi: 10.1186/1476-4598-10-2. Mol Cancer. 2011. PMID: 21211030 Free PMC article.
-
c-Ski inhibits the proliferation of vascular smooth muscle cells via suppressing Smad3 signaling but stimulating p38 pathway.Cell Signal. 2013 Jan;25(1):159-67. doi: 10.1016/j.cellsig.2012.09.001. Epub 2012 Sep 7. Cell Signal. 2013. PMID: 22986000
-
[Ski and SnoN: antagonistic proteins of TGFbeta signaling].Bull Cancer. 2000 Feb;87(2):135-7. Bull Cancer. 2000. PMID: 10705283 Review. French.
Cited by
-
Smad phosphoisoform signaling specificity: the right place at the right time.Carcinogenesis. 2011 Nov;32(11):1578-88. doi: 10.1093/carcin/bgr172. Epub 2011 Jul 27. Carcinogenesis. 2011. PMID: 21798854 Free PMC article. Review.
-
Smad phosphoisoform signals in acute and chronic liver injury: similarities and differences between epithelial and mesenchymal cells.Cell Tissue Res. 2012 Jan;347(1):225-43. doi: 10.1007/s00441-011-1178-6. Epub 2011 May 31. Cell Tissue Res. 2012. PMID: 21626291 Free PMC article. Review.
-
Next generation sequencing analysis of miRNAs: MiR-127-3p inhibits glioblastoma proliferation and activates TGF-β signaling by targeting SKI.OMICS. 2014 Mar;18(3):196-206. doi: 10.1089/omi.2013.0122. Epub 2014 Feb 11. OMICS. 2014. PMID: 24517116 Free PMC article.
-
SERPINE1 expression discriminates site-specific metastasis in human melanoma.Exp Dermatol. 2012 Jul;21(7):551-4. doi: 10.1111/j.1600-0625.2012.01523.x. Exp Dermatol. 2012. PMID: 22716255 Free PMC article.
-
Phospho-specific Smad3 signaling: impact on breast oncogenesis.Cell Cycle. 2012 Jul 1;11(13):2443-51. doi: 10.4161/cc.20546. Epub 2012 Jul 1. Cell Cycle. 2012. PMID: 22659843 Free PMC article. Review.
References
-
- Reed JA, Bales E, Xu W, Okan NA, Bandyopadhyay D, Medrano EE. Cytoplasmic localization of the oncogenic protein ski in human cutaneous melanomas in vivo: functional implications for transforming growth factor Beta signaling. Cancer Res. 2001;61:8074–8078. - PubMed
-
- Shinagawa T, Nomura T, Colmenares C, Ohira M, Nakagawara A, Ishii S. Increased susceptibility to tumorigenesis of ski-deficient heterozygous mice. Oncogene. 2001;20:8100–8108. - PubMed
-
- Le SE, Zhu Q, Wang L, Bandyopadhyay A, Javelaud D, Mauviel A, et al. Transforming growth factor-beta suppresses the ability of Ski to inhibit tumor metastasis by inducing its degradation. Cancer Res. 2008;68:3277–3285. - PubMed
-
- Nagano Y, Mavrakis KJ, Lee KL, Fujii T, Koinuma D, Sase H, et al. Arkadia induces degradation of SnoN and c-Ski to enhance transforming growth factor-beta signaling. J Biol Chem. 2007;282:20492–20501. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous