

Requirement for cyclin D3 in germinal center formation and function
- PMID: 20404856
- PMCID: PMC2997820
- DOI: 10.1038/cr.2010.55
Requirement for cyclin D3 in germinal center formation and function
Abstract
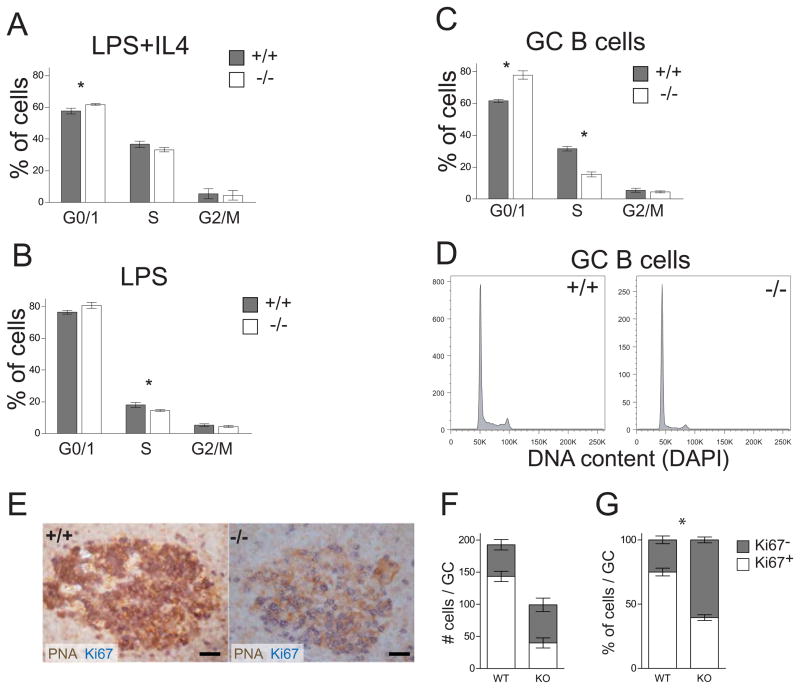
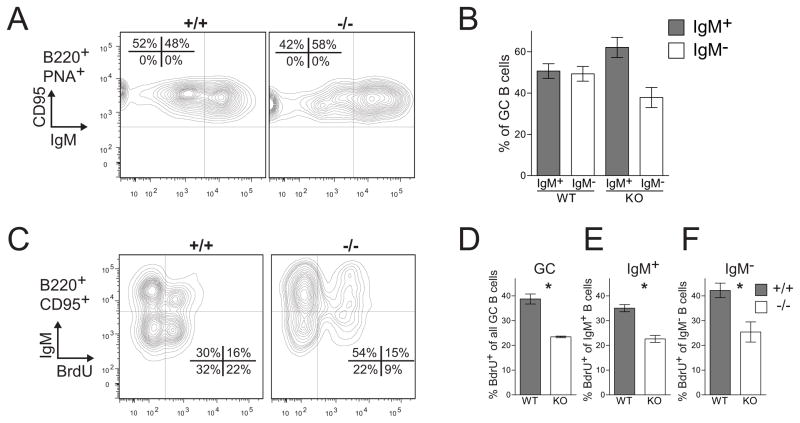
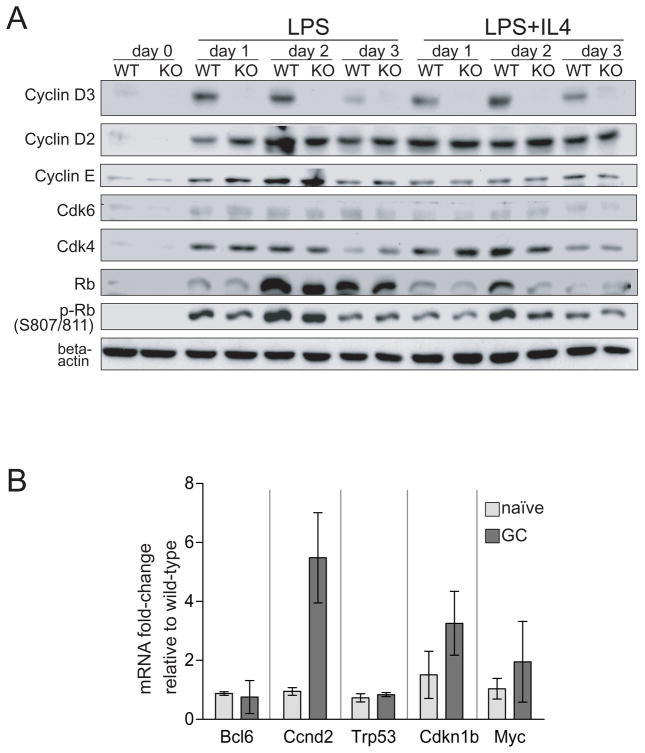
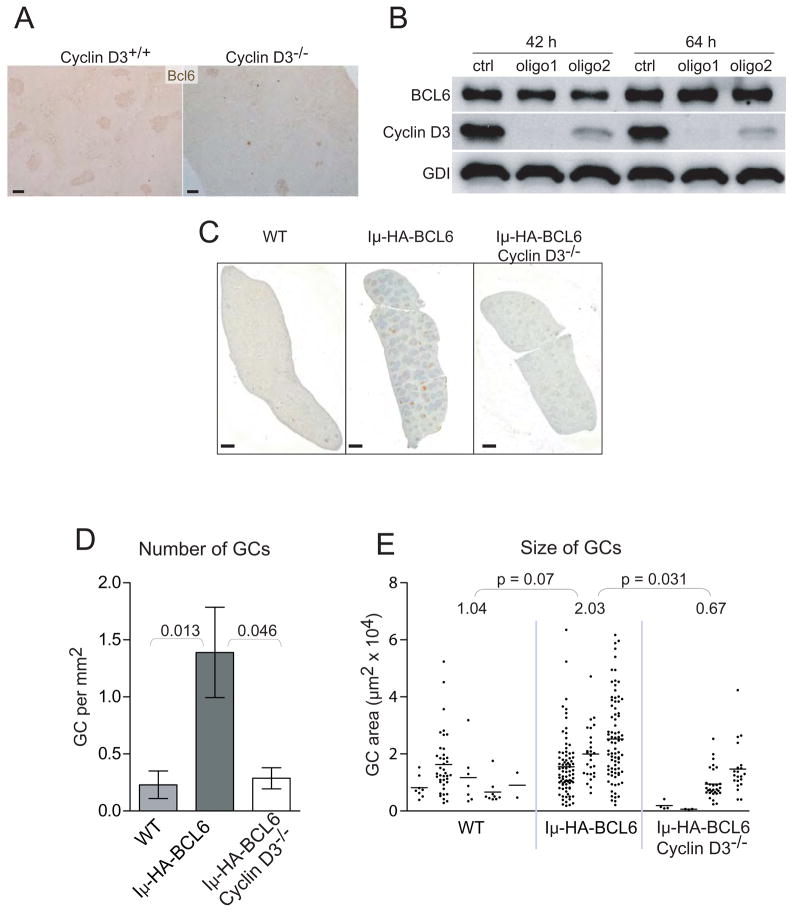
Germinal centers (GC) of secondary lymphoid tissues are critical to mounting a high-affinity humoral immune response. B cells within the GC undergo rapid clonal expansion and selection while diversifying their antibody genes. Although it is generally believed that GC B cells employ a unique proliferative program to accommodate these processes, little is known about how the GC-associated cell cycle is orchestrated. The D-type cyclins constitute an important component of the cell cycle engine that enables the cells to respond to physiological changes. Cell type- and developmental stage-specific roles of D-type cyclins have been described but the cyclin D requirement during GC reaction has not been addressed. In this study, we report that cyclin D3 is largely dispensable for proliferation and Ig class switching of in vitro activated B cells. In contrast, GC development in Ccnd3(-/-) mice is markedly impaired, as is the T cell-dependent antibody response. Within the GC, although both switched and unswitched B cells are affected by cyclin D3 inactivation, the IgM(-) pool is more severely reduced. Interestingly, despite a compensatory increase in cyclin D2 expression, a significant number of Ccnd3(-/-) GC B cells accumulate in quiescent G0 state. Lastly, although cyclin D3 inactivation did not disrupt BCL6 expression in GC B cells, it completely blocked the GC promoting effect of BCL6 overexpression, suggesting that cyclin D3 acts downstream of BCL6 to regulate GC formation. This is the first demonstration that cyclin D3 plays an important and unique role at the GC stage of B cell development.
Figures
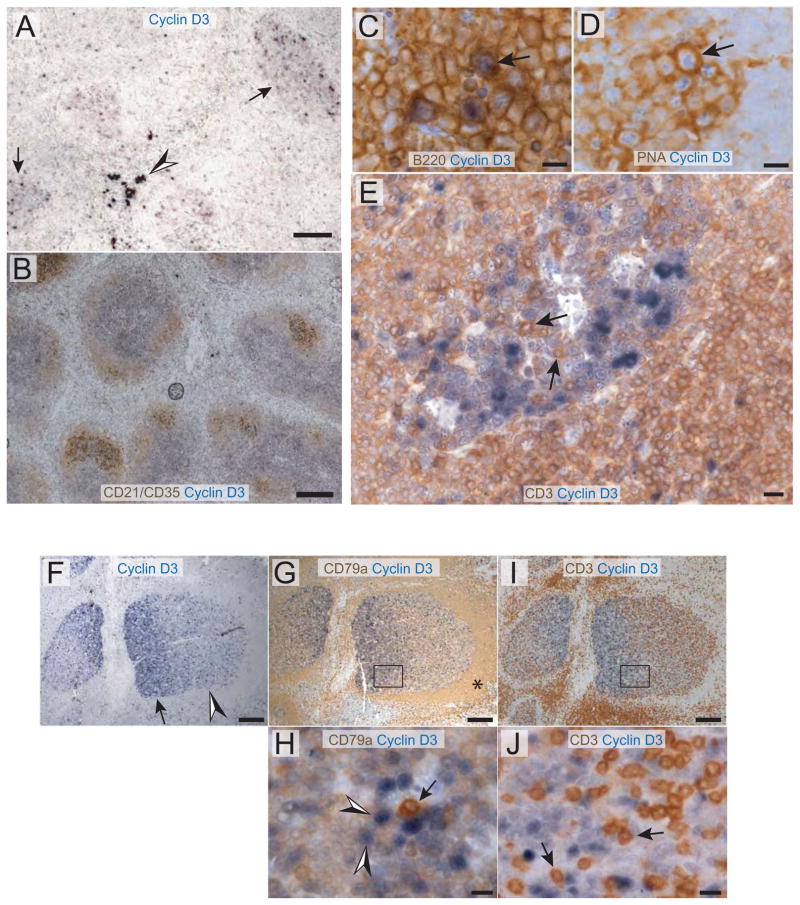
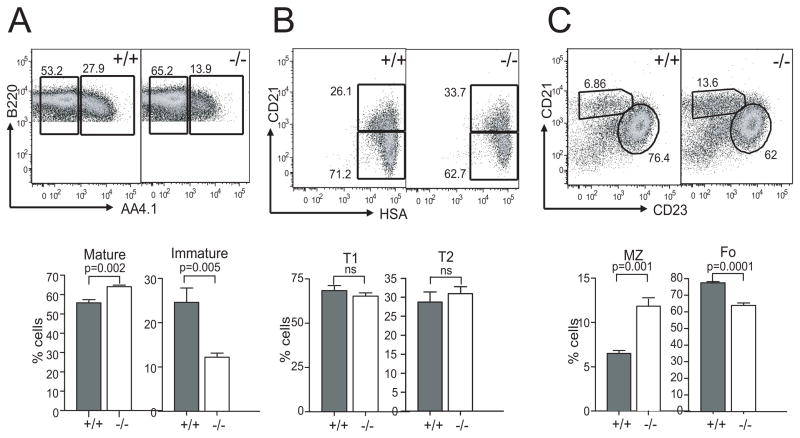
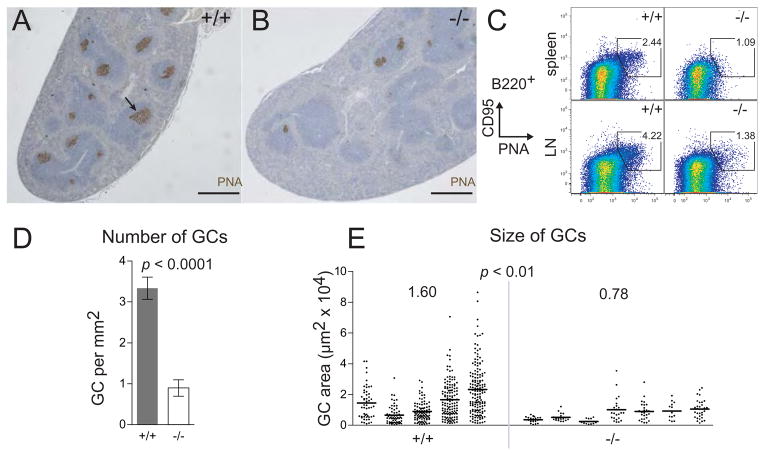
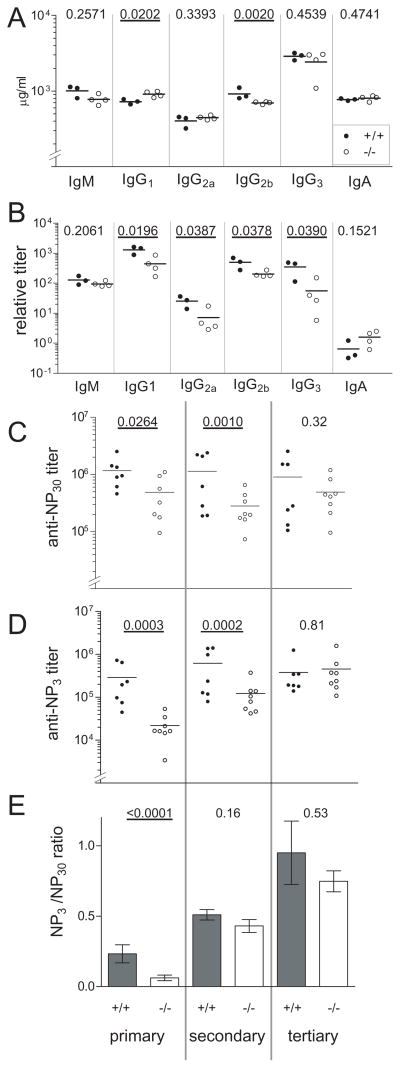
Similar articles
-
Cyclin D3 is selectively required for proliferative expansion of germinal center B cells.Mol Cell Biol. 2011 Jan;31(1):127-37. doi: 10.1128/MCB.00650-10. Epub 2010 Oct 18. Mol Cell Biol. 2011. PMID: 20956554 Free PMC article.
-
Cyclin D3 Governs Clonal Expansion of Dark Zone Germinal Center B Cells.Cell Rep. 2020 Nov 17;33(7):108403. doi: 10.1016/j.celrep.2020.108403. Cell Rep. 2020. PMID: 33207194 Free PMC article.
-
Cyclin D3 drives inertial cell cycling in dark zone germinal center B cells.J Exp Med. 2021 Apr 5;218(4):e20201699. doi: 10.1084/jem.20201699. J Exp Med. 2021. PMID: 33332554 Free PMC article.
-
Germinal center B-cells.Autoimmunity. 2012 Aug;45(5):333-47. doi: 10.3109/08916934.2012.665524. Epub 2012 Apr 2. Autoimmunity. 2012. PMID: 22390182 Review.
-
System-Level Scenarios for the Elucidation of T Cell-Mediated Germinal Center B Cell Differentiation.Front Immunol. 2021 Sep 20;12:734282. doi: 10.3389/fimmu.2021.734282. eCollection 2021. Front Immunol. 2021. PMID: 34616402 Free PMC article. Review.
Cited by
-
D-Type Cyclins in Development and Disease.Genes (Basel). 2023 Jul 14;14(7):1445. doi: 10.3390/genes14071445. Genes (Basel). 2023. PMID: 37510349 Free PMC article. Review.
-
Germinal Centre Shutdown.Front Immunol. 2021 Jul 7;12:705240. doi: 10.3389/fimmu.2021.705240. eCollection 2021. Front Immunol. 2021. PMID: 34305944 Free PMC article. Review.
-
Biology of Germinal Center B Cells Relating to Lymphomagenesis.Hemasphere. 2021 Jun 1;5(6):e582. doi: 10.1097/HS9.0000000000000582. eCollection 2021 Jun. Hemasphere. 2021. PMID: 34095765 Free PMC article. Review.
-
Translational repression of cyclin D3 by a stable G-quadruplex in its 5' UTR: implications for cell cycle regulation.RNA Biol. 2012 Aug;9(8):1099-109. doi: 10.4161/rna.21210. Epub 2012 Aug 1. RNA Biol. 2012. PMID: 22858673 Free PMC article.
-
Synergy between PI3K signaling and MYC in Burkitt lymphomagenesis.Cancer Cell. 2012 Aug 14;22(2):167-79. doi: 10.1016/j.ccr.2012.06.012. Cancer Cell. 2012. PMID: 22897848 Free PMC article.
References
-
- Peled JU, Kuang FL, Iglesias-Ussel MD, et al. The biochemistry of somatic hypermutation. Annu Rev Immunol. 2008;26:481–511. - PubMed
-
- Wang Y, Carter RH. CD19 regulates B cell maturation, proliferation, and positive selection in the FDC zone of murine splenic germinal centers. Immunity. 2005;22 (6):749–761. - PubMed
-
- Klein U, Dalla-Favera R. Germinal centres: role in B-cell physiology and malignancy. Nat Rev Immunol. 2008;8 (1):22–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K08 CA127353/CA/NCI NIH HHS/United States
- R01 AR049126/AR/NIAMS NIH HHS/United States
- R01 CA085573/CA/NCI NIH HHS/United States
- R01 CA108420/CA/NCI NIH HHS/United States
- CA102705/CA/NCI NIH HHS/United States
- T32GM007288/GM/NIGMS NIH HHS/United States
- P01 AI051392/AI/NIAID NIH HHS/United States
- P01 CA109901/CA/NCI NIH HHS/United States
- R01 CA72649/CA/NCI NIH HHS/United States
- AI051392/AI/NIAID NIH HHS/United States
- R01 CA85573/CA/NCI NIH HHS/United States
- R01 CA102705/CA/NCI NIH HHS/United States
- T32 GM007288/GM/NIGMS NIH HHS/United States
- R01 CA072649/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
