

Dysferlin interacts with tubulin and microtubules in mouse skeletal muscle
- PMID: 20405035
- PMCID: PMC2853571
- DOI: 10.1371/journal.pone.0010122
Dysferlin interacts with tubulin and microtubules in mouse skeletal muscle
Abstract
Dysferlin is a type II transmembrane protein implicated in surface membrane repair in muscle. Mutations in dysferlin lead to limb girdle muscular dystrophy 2B, Miyoshi Myopathy and distal anterior compartment myopathy. Dysferlin's mode of action is not well understood and only a few protein binding partners have thus far been identified. Using affinity purification followed by liquid chromatography/mass spectrometry, we identified alpha-tubulin as a novel binding partner for dysferlin. The association between dysferlin and alpha-tubulin, as well as between dysferlin and microtubules, was confirmed in vitro by glutathione S-transferase pulldown and microtubule binding assays. These interactions were confirmed in vivo by co-immunoprecipitation. Confocal microscopy revealed that dysferlin and alpha-tubulin co-localized in the perinuclear region and in vesicular structures in myoblasts, and along thin longitudinal structures reminiscent of microtubules in myotubes. We mapped dysferlin's alpha-tubulin-binding region to its C2A and C2B domains. Modulation of calcium levels did not affect dysferlin binding to alpha-tubulin, suggesting that this interaction is calcium-independent. Our studies identified a new binding partner for dysferlin and suggest a role for microtubules in dysferlin trafficking to the sarcolemma.
Conflict of interest statement
Figures
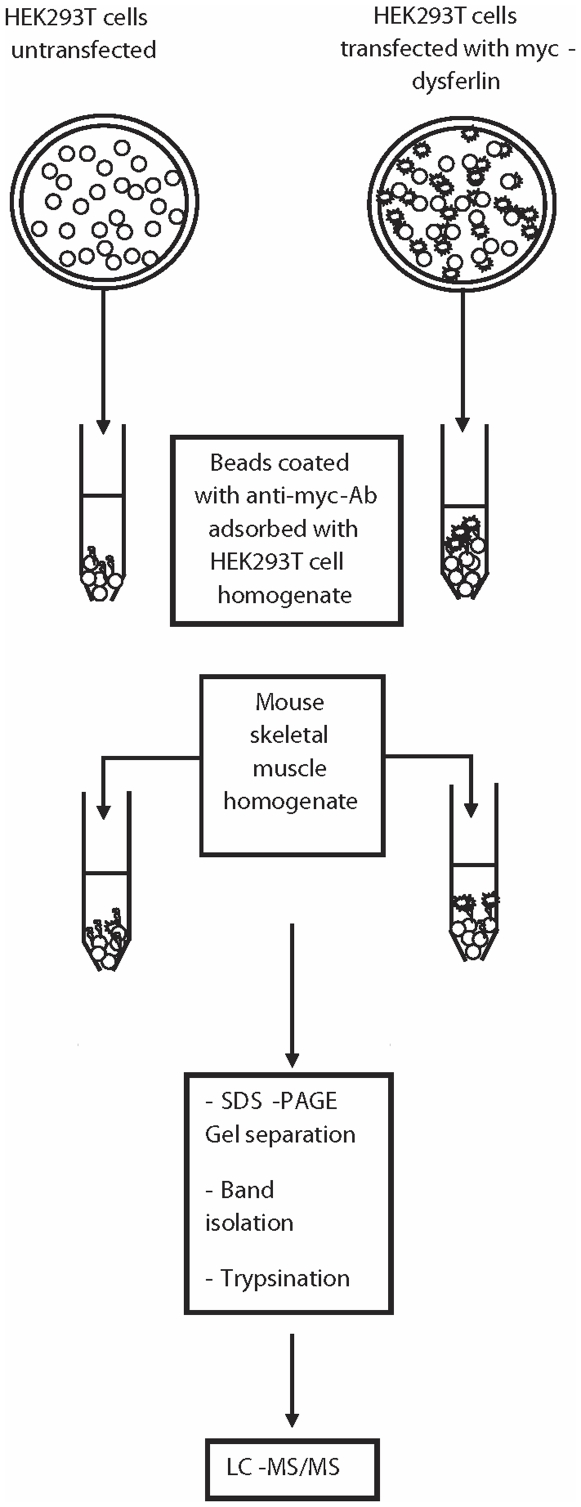
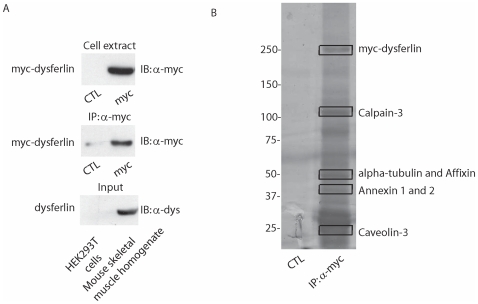
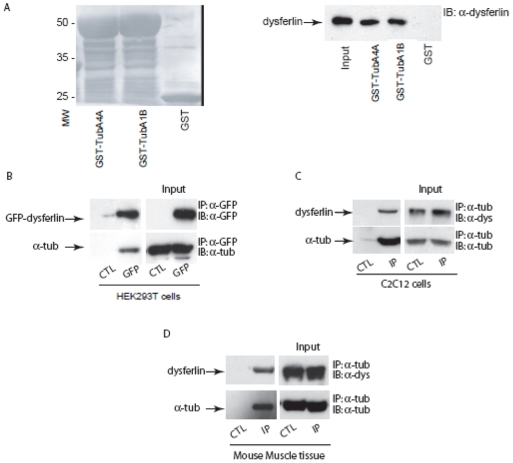
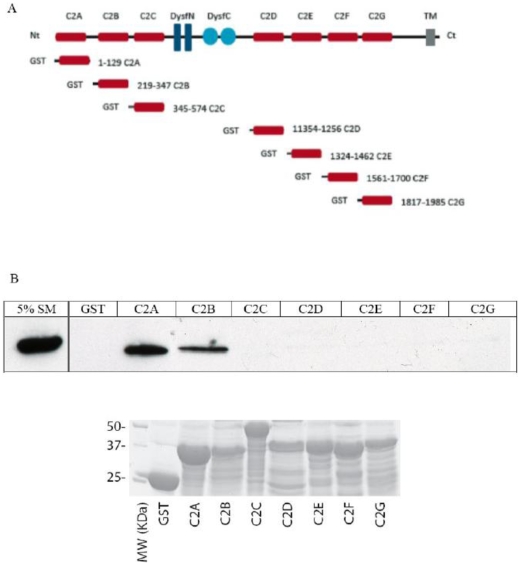
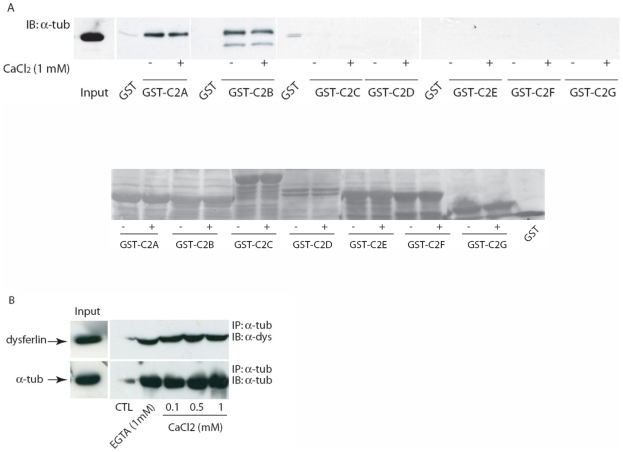

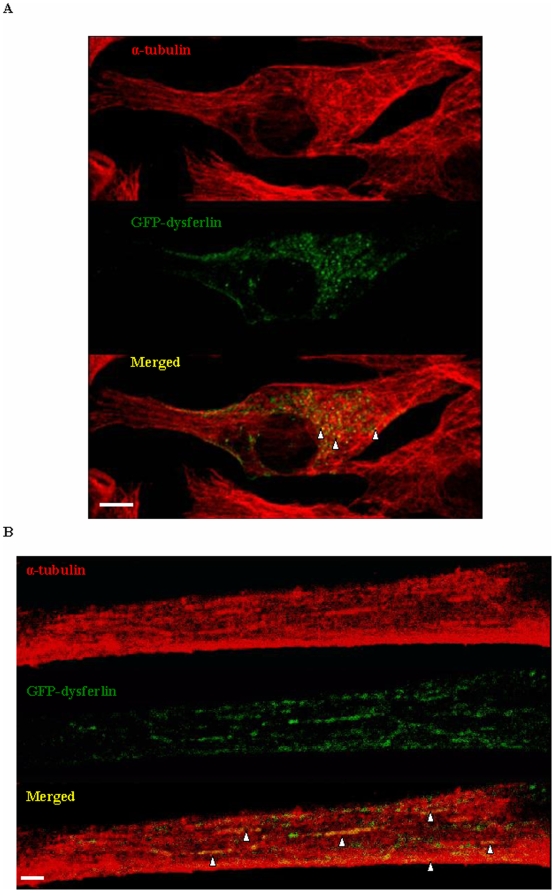
References
-
- Bashir R, Britton S, Strachan T, Keers S, Vafiadaki E, et al. A gene related to Caenorhabditis elegans spermatogenesis factor fer-1 is mutated in limb-girdle muscular dystrophy type 2B. Nat Genet. 1998;20:37–42. - PubMed
-
- Liu J, Aoki M, Illa I, Wu C, Fardeau M, et al. Dysferlin, a novel skeletal muscle gene, is mutated in Miyoshi myopathy and limb girdle muscular dystrophy. Nat Genet. 1998;20:31–36. - PubMed
-
- Illa I, Serrano-Munuera C, Gallardo E, Lasa A, Rojas-Garcia R, et al. Distal anterior compartment myopathy: a dysferlin mutation causing a new muscular dystrophy phenotype. Ann Neurol. 2001;49:130–134. - PubMed
-
- Therrien C, Dodig D, Karpati G, Sinnreich M. Mutation impact on dysferlin inferred from database analysis and computer-based structural predictions. Journal of the Neurological Sciences. 2006;250:71–78. - PubMed
-
- Vandre DD, Ackerman WEt, Kniss DA, Tewari AK, Mori M, et al. Dysferlin is expressed in human placenta but does not associate with caveolin. Biol Reprod. 2007;77:533–542. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
