

Hypothalamic pituitary adrenal axis responses to low-intensity stressors are reduced after voluntary wheel running in rats
- PMID: 20406350
- PMCID: PMC4469265
- DOI: 10.1111/j.1365-2826.2010.02007.x
Hypothalamic pituitary adrenal axis responses to low-intensity stressors are reduced after voluntary wheel running in rats
Abstract
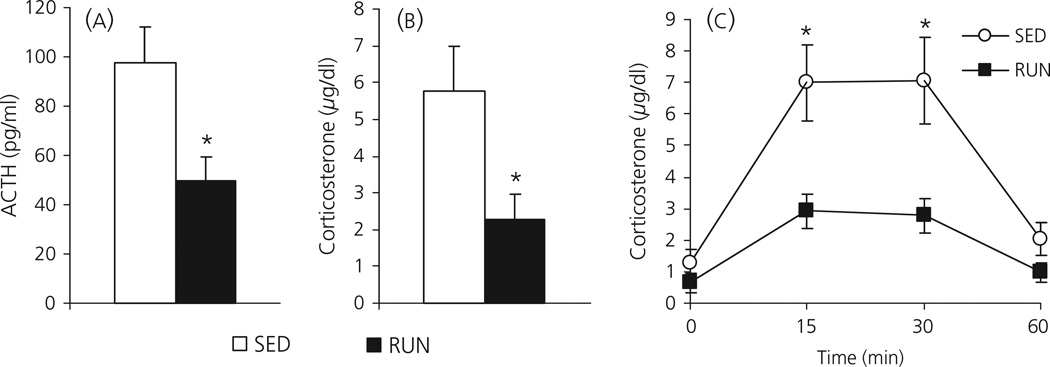
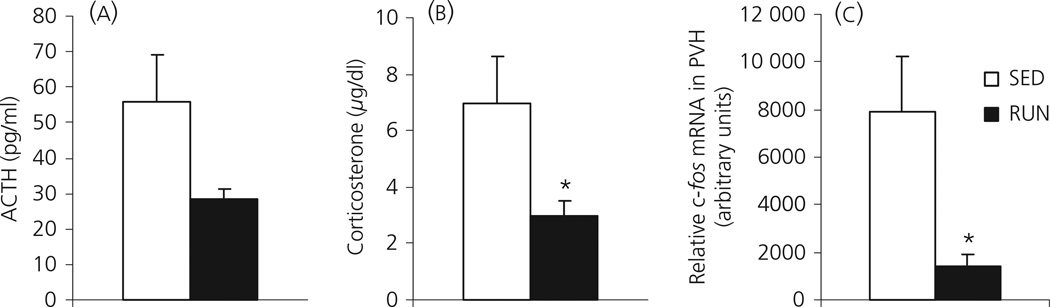
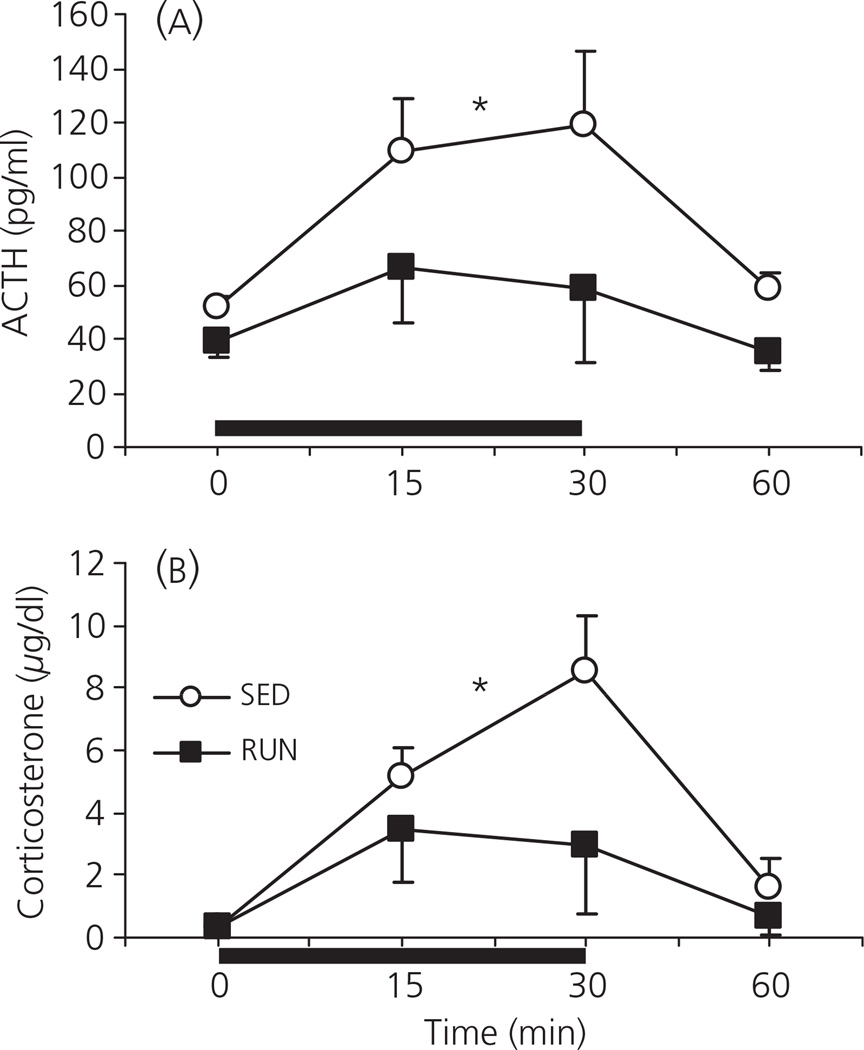
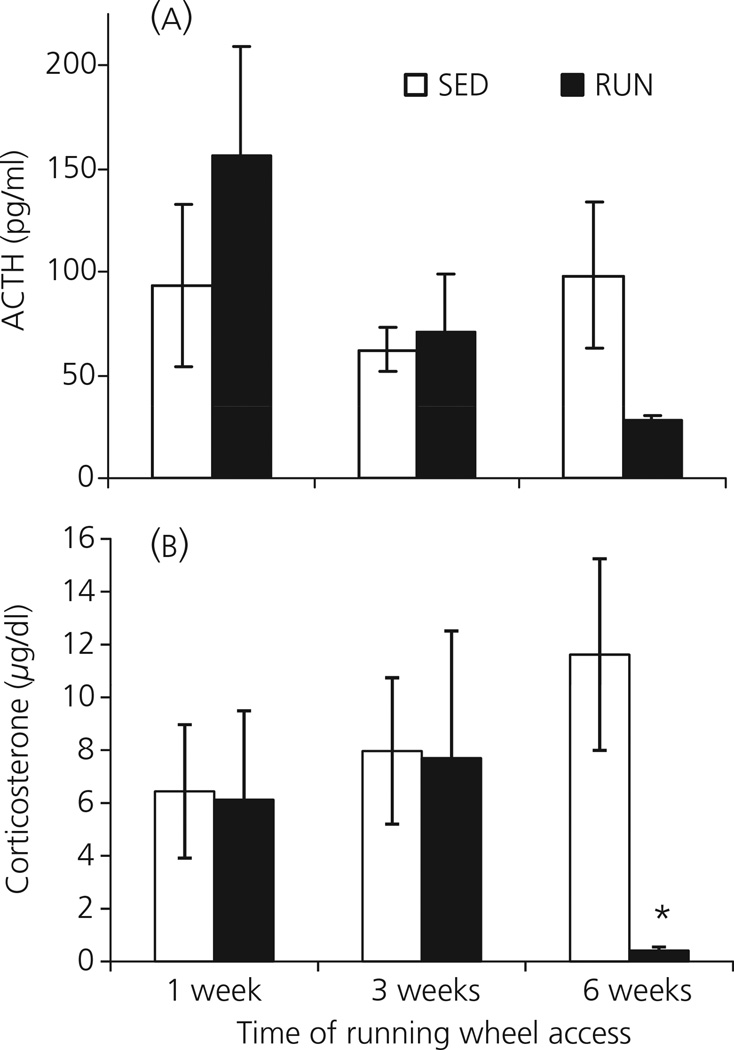
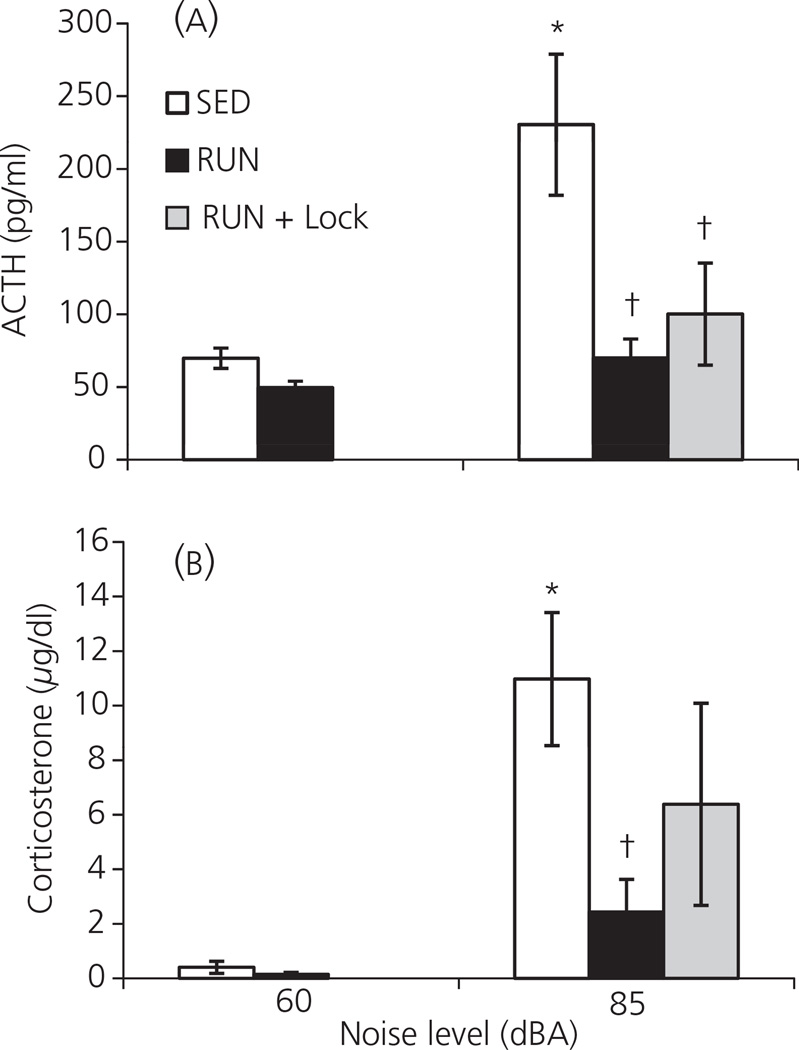
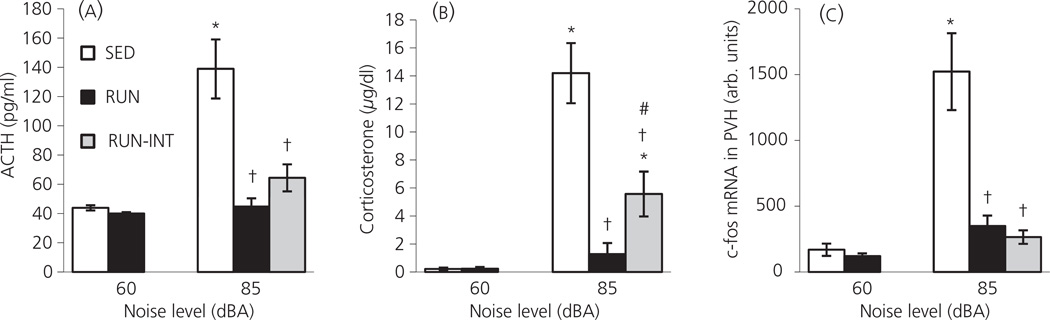
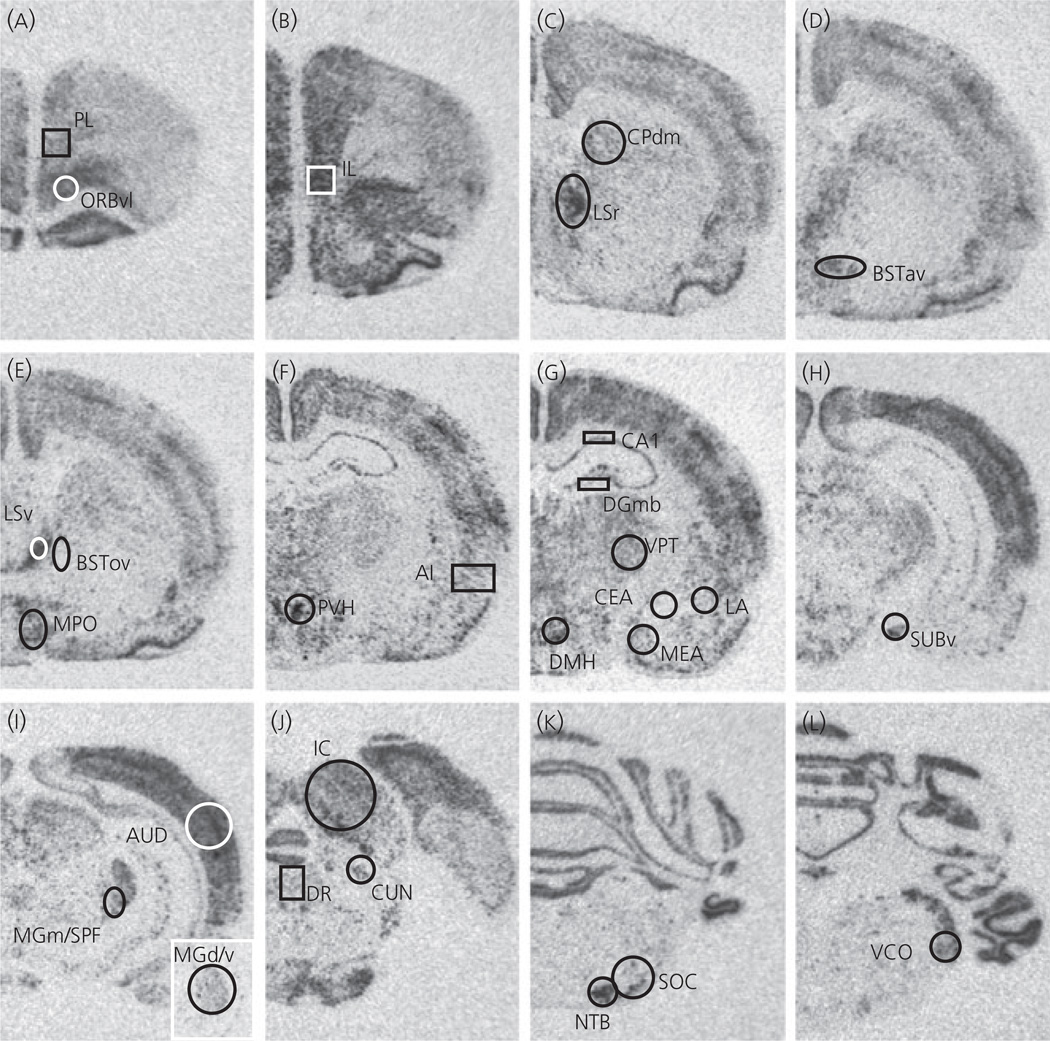
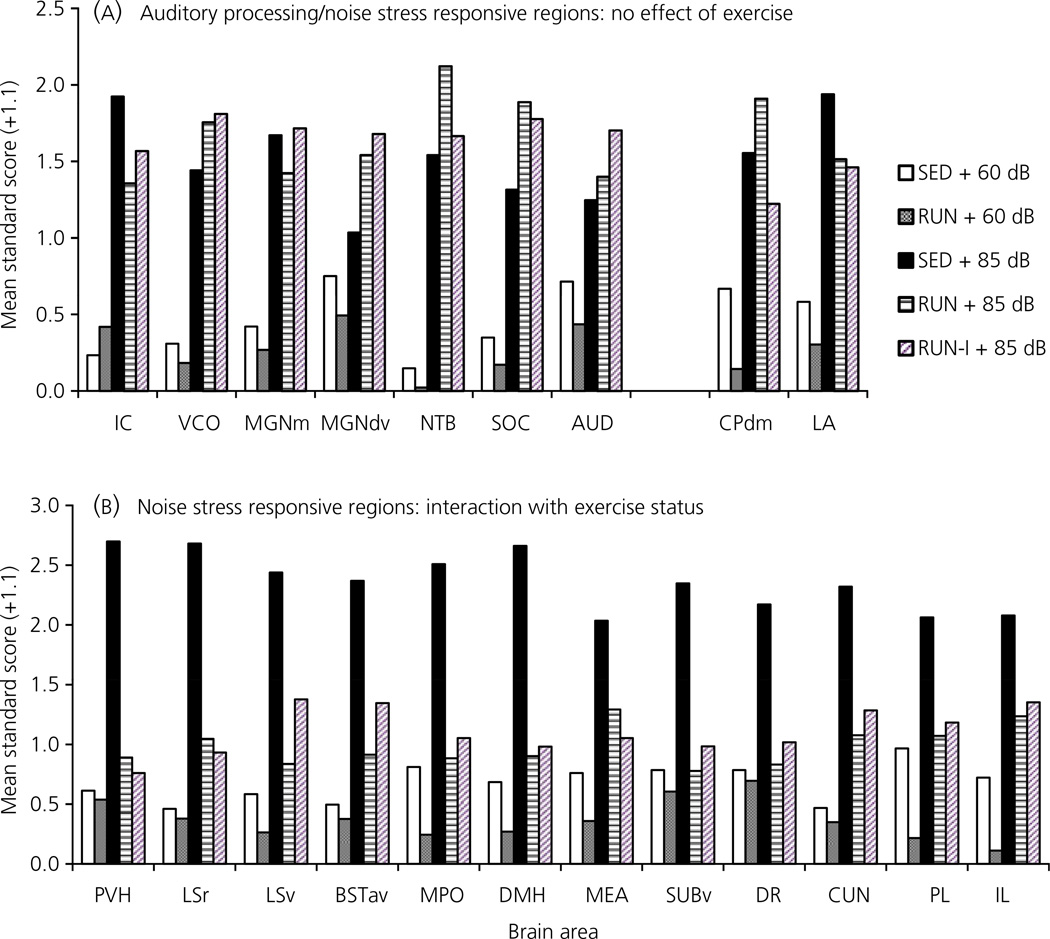
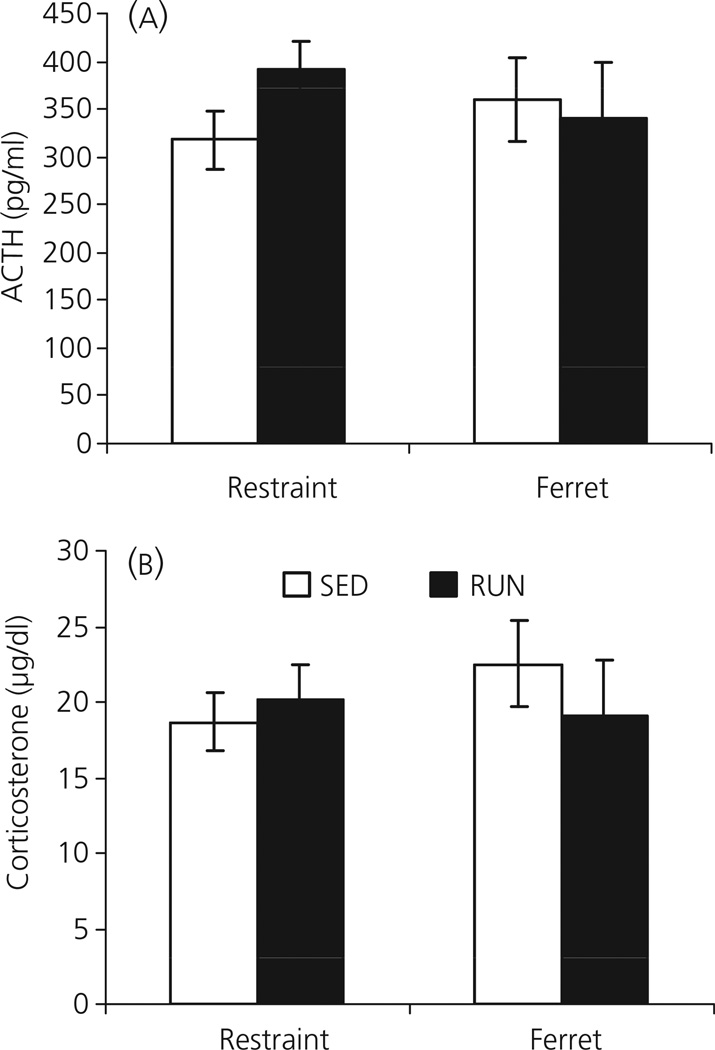
Regular physical exercise is beneficial for both physical and mental health. By contrast, stress is associated with deleterious effects on health and there is growing evidence that regular physical exercise counteracts some of the effects of stress. However, most previous studies have suggested that prior exercise does not alter the acute hypothalamic pituitary adrenal (HPA) axis responses to stress. The present series of studies provides evidence that in rats, 6 weeks (but not 1 or 3 weeks) of voluntary wheel running reduces the HPA axis responses to lower-intensity stressors such as an i.p. saline injection, exposure to a novel environment or exposure to moderate intensity noise, but not to more intense stressors such as predator odour exposure or restraint. Daily exercise does not appear to be necessary for the reduction in HPA axis responses, with intermittent access (24 h out of each 72-h period) to a running wheel for 6 weeks, resulting in similar decrements in adrenocorticotrophic hormone and corticosterone release in response to 85 dBA noise exposure. Data from in situ hybridisation for c-fos mRNA are consistent with the hypothesis that voluntary exercise results in a decrease in HPA axis responsiveness to a low-intensity stressor at a central level, with no changes in primary sensory processing. Together, these data suggest that 6 weeks of daily or intermittent exercise constrains the HPA axis response to mild, but not more intense stressors, and that this regulation may be mediated at a central level beyond the primary sensory input.
Figures
Similar articles
-
Stressor-specific effects of sex on HPA axis hormones and activation of stress-related neurocircuitry.Stress. 2013 Nov;16(6):664-77. doi: 10.3109/10253890.2013.840282. Epub 2013 Oct 1. Stress. 2013. PMID: 23992519 Free PMC article.
-
Voluntary wheel running modulates glutamate receptor subunit gene expression and stress hormone release in Lewis rats.Psychoneuroendocrinology. 2003 Jul;28(5):702-14. doi: 10.1016/s0306-4530(02)00062-8. Psychoneuroendocrinology. 2003. PMID: 12727136
-
Voluntary exercise impacts on the rat hypothalamic-pituitary-adrenocortical axis mainly at the adrenal level.Neuroendocrinology. 2007;86(1):26-37. doi: 10.1159/000104770. Epub 2007 Jun 25. Neuroendocrinology. 2007. PMID: 17595533
-
A single exposure to severe stressors causes long-term desensitisation of the physiological response to the homotypic stressor.Stress. 2004 Sep;7(3):157-72. doi: 10.1080/10253890400010721. Stress. 2004. PMID: 15764013 Review.
-
Central mechanisms of HPA axis regulation by voluntary exercise.Neuromolecular Med. 2008;10(2):118-27. doi: 10.1007/s12017-008-8027-0. Epub 2008 Feb 14. Neuromolecular Med. 2008. PMID: 18273712 Free PMC article. Review.
Cited by
-
Effects of physical exercise on central nervous system functions: a review of brain region specific adaptations.J Mol Psychiatry. 2015 Apr 18;3(1):3. doi: 10.1186/s40303-015-0010-8. eCollection 2015. J Mol Psychiatry. 2015. PMID: 26064521 Free PMC article.
-
Exercise, stress resistance, and central serotonergic systems.Exerc Sport Sci Rev. 2011 Jul;39(3):140-9. doi: 10.1097/JES.0b013e31821f7e45. Exerc Sport Sci Rev. 2011. PMID: 21508844 Free PMC article. Review.
-
Redox Regulatory Changes of Circadian Rhythm by the Environmental Risk Factors Traffic Noise and Air Pollution.Antioxid Redox Signal. 2022 Oct;37(10-12):679-703. doi: 10.1089/ars.2021.0272. Epub 2022 Apr 18. Antioxid Redox Signal. 2022. PMID: 35088601 Free PMC article. Review.
-
Effects of Exercise on Depression and Anxiety in Lung Cancer Survivors: A Systematic Review and Meta-Analysis of Randomized Controlled Trials.Curr Oncol. 2025 May 25;32(6):304. doi: 10.3390/curroncol32060304. Curr Oncol. 2025. PMID: 40558247 Free PMC article. Review.
-
Stressor-specific effects of sex on HPA axis hormones and activation of stress-related neurocircuitry.Stress. 2013 Nov;16(6):664-77. doi: 10.3109/10253890.2013.840282. Epub 2013 Oct 1. Stress. 2013. PMID: 23992519 Free PMC article.
References
-
- Cotman CW, Berchtold NC, Christie LA. Exercise builds brain health: key roles of growth factor cascades and inflammation. Trends Neurosci. 2007;30:464–472. - PubMed
-
- Dishman RK, Berthoud HR, Booth FW, Cotman CW, Edgerton VR, Fleshner MR, Gandevia SC, Gomez-Pinilla F, Greenwood BN, Hillman CH, Kramer AF, Levin BE, Moran TH, Russo-Neustadt AA, Salamone JD, Van Hoomissen JD, Wade CE, York DA, Zigmond MJ. Neurobiology of exercise. Obesity (Silver Spring) 2006;14:345–356. - PubMed
-
- Fleshner M. Physical activity and stress resistance: sympathetic nervous system adaptations prevent stress-induced immunosuppression. Exerc Sport Sci Rev. 2005;33:120–126. - PubMed
-
- Greenwood BN, Fleshner M. Exercise, learned helplessness, and the stress-resistant brain. Neuromolecular Med. 2008;10:81–98. - PubMed
-
- Moraska A, Fleshner M. Voluntary physical activity prevents stress-induced behavioral depression and anti-KLH antibody suppression. Am J Physiol Regul Integr Comp Physiol. 2001;281:R484–R489. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
