

Regulation of MCP-1 chemokine transcription by p53
- PMID: 20406462
- PMCID: PMC2864217
- DOI: 10.1186/1476-4598-9-82
Regulation of MCP-1 chemokine transcription by p53
Abstract
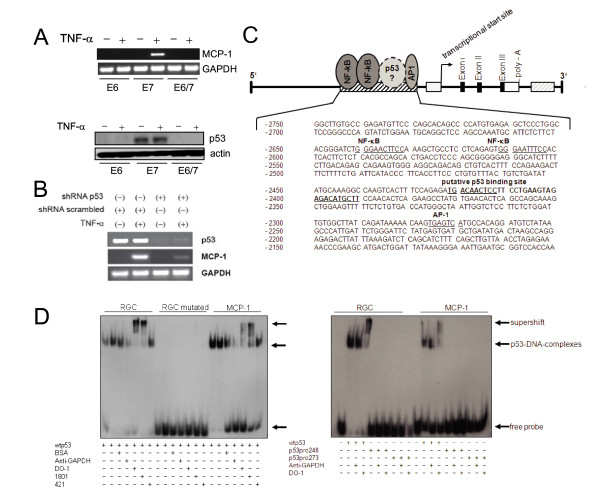
Background: Our previous studies showed that the expression of the monocyte-chemoattractant protein (MCP)-1, a chemokine, which triggers the infiltration and activation of cells of the monocyte-macrophage lineage, is abrogated in human papillomavirus (HPV)-positive premalignant and malignant cells. In silico analysis of the MCP-1 upstream region proposed a putative p53 binding side about 2.5 kb upstream of the transcriptional start. The aim of this study is to monitor a physiological role of p53 in this process.
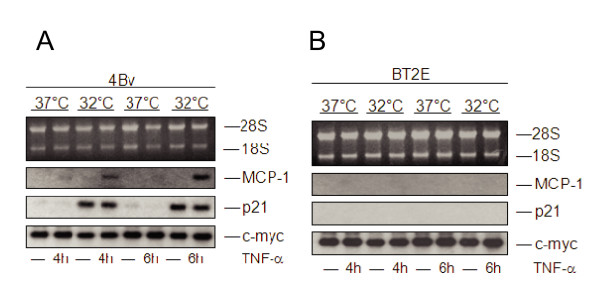
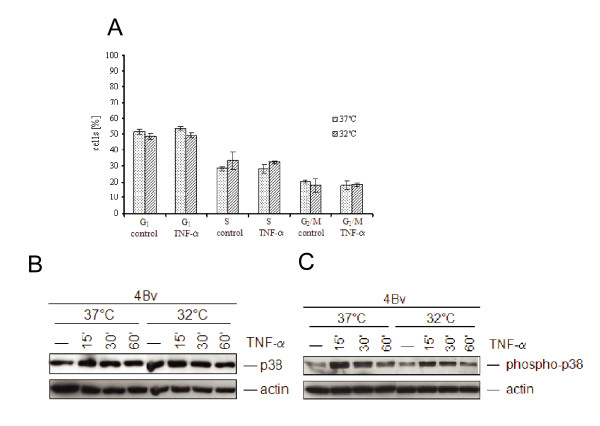
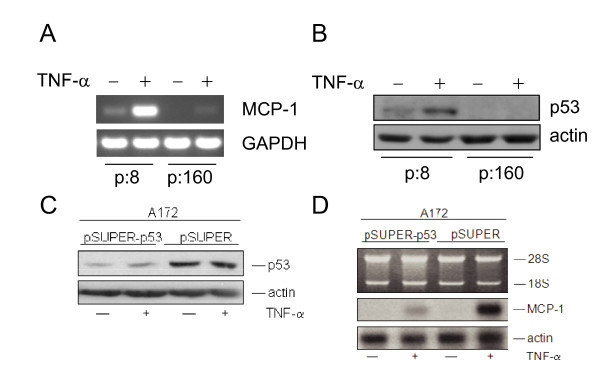
Results: The proposed p53 binding side could be confirmed in vitro by electrophoretic-mobility-shift assays and in vivo by chromatin immunoprecipitation. Moreover, the availability of p53 is apparently important for chemokine regulation, since TNF-alpha can induce MCP-1 only in human keratinocytes expressing the viral oncoprotein E7, but not in HPV16 E6 positive cells, where p53 becomes degraded. A general physiological role of p53 in MCP-1 regulation was further substantiated in HPV-negative cells harboring a temperature-sensitive mutant of p53 and in Li-Fraumeni cells, carrying a germ-line mutation of p53. In both cases, non-functional p53 leads to diminished MCP-1 transcription upon TNF-alpha treatment. In addition, siRNA directed against p53 decreased MCP-1 transcription after TNF-alpha addition, directly confirming a crosstalk between p53 and MCP-1.
Conclusion: These data support the concept that p53 inactivation during carcinogenesis also affects immune surveillance by interfering with chemokine expression and in turn communication with cells of the immunological compartment.
Figures

Similar articles
-
A role for NF-kappa B binding motifs in the differential induction of chemokine gene expression in human corneal epithelial cells.Invest Ophthalmol Vis Sci. 2004 Jul;45(7):2299-305. doi: 10.1167/iovs.03-0367. Invest Ophthalmol Vis Sci. 2004. PMID: 15223809
-
Transcription factor activating protein-2beta: a positive regulator of monocyte chemoattractant protein-1 gene expression.Endocrinology. 2009 Apr;150(4):1654-61. doi: 10.1210/en.2008-1361. Epub 2008 Nov 20. Endocrinology. 2009. PMID: 19022887
-
Molecular analysis of the inhibition of monocyte chemoattractant protein-1 gene expression by estrogens and xenoestrogens in MCF-7 cells.Endocrinology. 2000 Jan;141(1):50-9. doi: 10.1210/endo.141.1.7233. Endocrinology. 2000. PMID: 10614622
-
Dexamethasone regulates AP-1 to repress TNF-alpha induced MCP-1 production in human glomerular endothelial cells.Nephrol Dial Transplant. 2004 Feb;19(2):312-9. doi: 10.1093/ndt/gfg583. Nephrol Dial Transplant. 2004. PMID: 14736953
-
Induction of IL-8 and monoclyte chemoattractant protein-1 by doxorubicin in human small cell lung carcinoma cells.Int J Cancer. 2003 Jan 20;103(3):380-6. doi: 10.1002/ijc.10842. Int J Cancer. 2003. PMID: 12471621
Cited by
-
Triggering Degradation of Host Cellular Proteins for Robust Propagation of Influenza Viruses.Int J Mol Sci. 2024 Apr 25;25(9):4677. doi: 10.3390/ijms25094677. Int J Mol Sci. 2024. PMID: 38731896 Free PMC article. Review.
-
Human papillomavirus and cervical cancer: an insight highlighting pathogenesis and targeting strategies.Virusdisease. 2022 Jun;33(2):132-154. doi: 10.1007/s13337-022-00768-w. Epub 2022 May 30. Virusdisease. 2022. PMID: 35991700 Free PMC article. Review.
-
p53 serves as a host antiviral factor that enhances innate and adaptive immune responses to influenza A virus.J Immunol. 2011 Dec 15;187(12):6428-36. doi: 10.4049/jimmunol.1101459. Epub 2011 Nov 21. J Immunol. 2011. PMID: 22105999 Free PMC article.
-
Epidemiology, Molecular Pathogenesis, Immuno-Pathogenesis, Immune Escape Mechanisms and Vaccine Evaluation for HPV-Associated Carcinogenesis.Pathogens. 2023 Nov 23;12(12):1380. doi: 10.3390/pathogens12121380. Pathogens. 2023. PMID: 38133265 Free PMC article. Review.
-
HPV and the Risk of HIV Acquisition in Women.Front Cell Infect Microbiol. 2022 Feb 10;12:814948. doi: 10.3389/fcimb.2022.814948. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35223546 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
