

Apobec-1 complementation factor modulates liver regeneration by post-transcriptional regulation of interleukin-6 mRNA stability
- PMID: 20406809
- PMCID: PMC2885197
- DOI: 10.1074/jbc.M110.115147
Apobec-1 complementation factor modulates liver regeneration by post-transcriptional regulation of interleukin-6 mRNA stability
Abstract
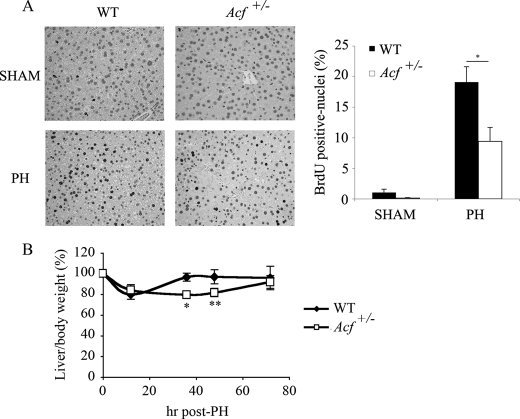
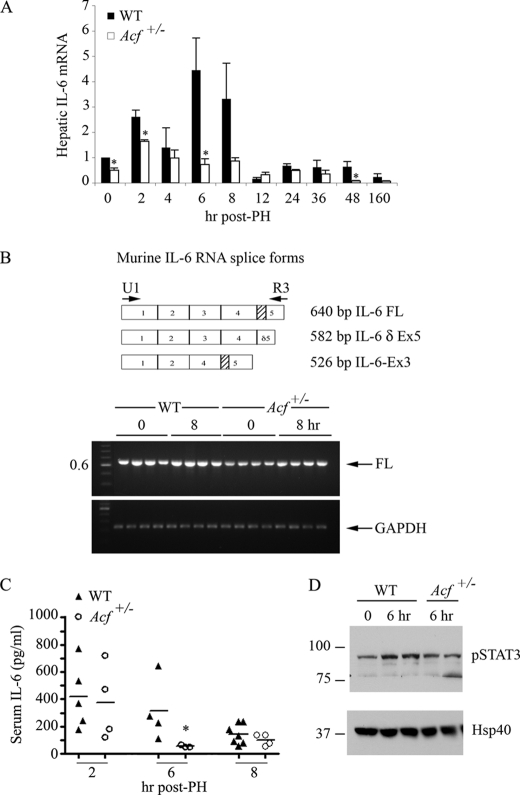
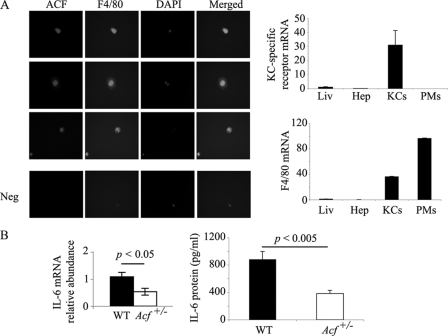
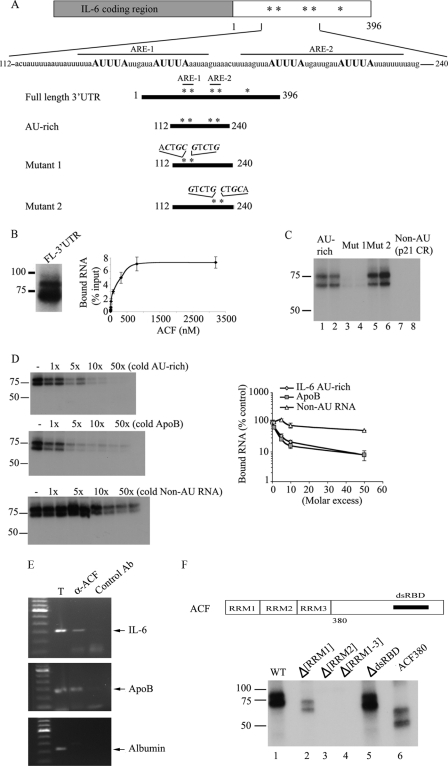
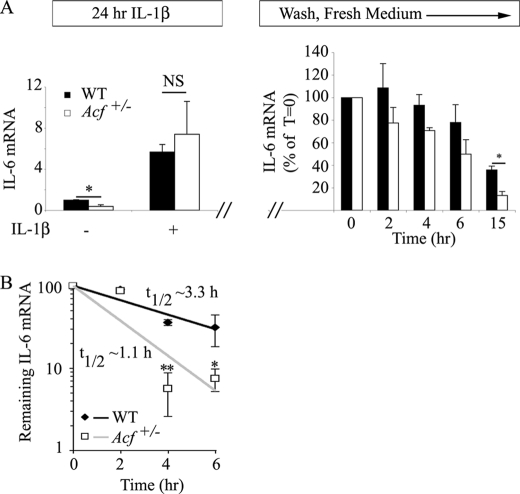
Apobec-1 complementation factor (ACF) is the RNA binding subunit of a core complex that mediates C to U RNA editing of apolipoprotein B (apoB) mRNA. Targeted deletion of the murine Acf gene is early embryonic lethal and Acf(-/-) blastocysts fail to implant and proliferate, suggesting that ACF plays a key role in cell growth and differentiation. Here we demonstrate that heterozygous Acf(+/-) mice exhibit decreased proliferation and impaired liver mass restitution following partial hepatectomy (PH). To pursue the mechanism of impaired liver regeneration we examined activation of interleukin-6 (IL-6) a key cytokine required for induction of hepatocyte proliferation following PH. Peak induction of hepatic IL-6 mRNA abundance post PH was attenuated >80% in heterozygous Acf(+/-) mice, along with decreased serum IL-6 levels. IL-6 secretion from isolated Kupffer cells (KC) was 2-fold greater in wild-type compared with heterozygous Acf(+/-) mice. Recombinant ACF bound an AU-rich region in the IL-6 3'-untranslated region with high affinity and IL-6 mRNA half-life was significantly shorter in KC isolated from Acf(+/-) mice compared with wild-type controls. These findings suggest that ACF regulates liver regeneration following PH at least in part by controlling the stability of IL-6 mRNA. The results further suggest a new RNA target and an unanticipated physiological function for ACF beyond apoB RNA editing.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
