

Protein structural dynamics in solution unveiled via 100-ps time-resolved x-ray scattering
- PMID: 20406909
- PMCID: PMC2867760
- DOI: 10.1073/pnas.1002951107
Protein structural dynamics in solution unveiled via 100-ps time-resolved x-ray scattering
Abstract
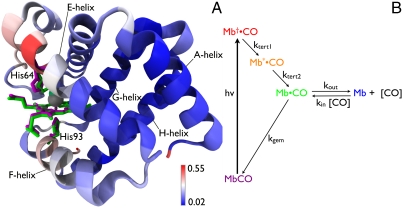
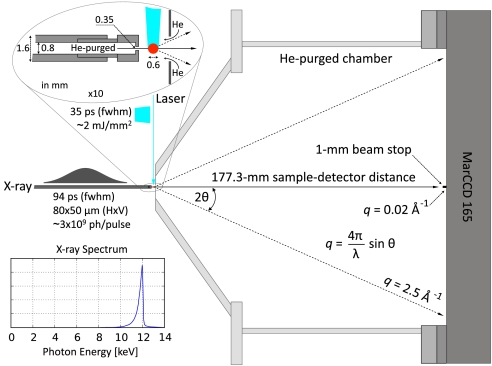
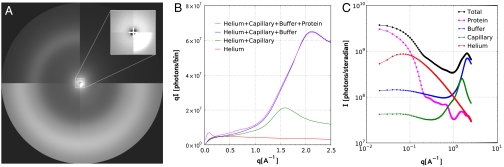
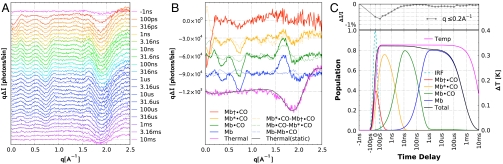
We have developed a time-resolved x-ray scattering diffractometer capable of probing structural dynamics of proteins in solution with 100-ps time resolution. This diffractometer, developed on the ID14B BioCARS (Consortium for Advanced Radiation Sources) beamline at the Advanced Photon Source, records x-ray scattering snapshots over a broad range of q spanning 0.02-2.5 A(-1), thereby providing simultaneous coverage of the small-angle x-ray scattering (SAXS) and wide-angle x-ray scattering (WAXS) regions. To demonstrate its capabilities, we have tracked structural changes in myoglobin as it undergoes a photolysis-induced transition from its carbon monoxy form (MbCO) to its deoxy form (Mb). Though the differences between the MbCO and Mb crystal structures are small (rmsd < 0.2 A), time-resolved x-ray scattering differences recorded over 8 decades of time from 100 ps to 10 ms are rich in structure, illustrating the sensitivity of this technique. A strong, negative-going feature in the SAXS region appears promptly and corresponds to a sudden > 22 A(3) volume expansion of the protein. The ensuing conformational relaxation causes the protein to contract to a volume approximately 2 A(3) larger than MbCO within approximately 10 ns. On the timescale for CO escape from the primary docking site, another change in the SAXS/WAXS fingerprint appears, demonstrating sensitivity to the location of the dissociated CO. Global analysis of the SAXS/WAXS patterns recovered time-independent scattering fingerprints for four intermediate states of Mb. These SAXS/WAXS fingerprints provide stringent constraints for putative models of conformational states and structural transitions between them.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Schotte F, et al. Watching a protein as it functions with 150-ps time-resolved x-ray crystallography. Science. 2003;300:1944–1947. - PubMed
-
- Schotte F, Soman J, Olson JS, Wulff M, Anfinrud PA. Picosecond time-resolved x-ray crystallography: Probing protein function in real time. J Struct Biol. 2004;147:235–246. - PubMed
-
- Haurowitz F. The Equilibrium between hemoglobin and oxygen. H-S Z Physiol Chem. 1938;254:266–272.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
