

The Drosophila Claudin Kune-kune is required for septate junction organization and tracheal tube size control
- PMID: 20407131
- PMCID: PMC2907205
- DOI: 10.1534/genetics.110.114959
The Drosophila Claudin Kune-kune is required for septate junction organization and tracheal tube size control
Abstract
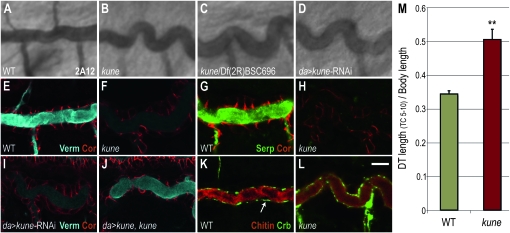
The vertebrate tight junction is a critical claudin-based cell-cell junction that functions to prevent free paracellular diffusion between epithelial cells. In Drosophila, this barrier is provided by the septate junction, which, despite being ultrastructurally distinct from the vertebrate tight junction, also contains the claudin-family proteins Megatrachea and Sinuous. Here we identify a third Drosophila claudin, Kune-kune, that localizes to septate junctions and is required for junction organization and paracellular barrier function, but not for apical-basal polarity. In the tracheal system, septate junctions have a barrier-independent function that promotes lumenal secretion of Vermiform and Serpentine, extracellular matrix modifier proteins that are required to restrict tube length. As with Sinuous and Megatrachea, loss of Kune-kune prevents this secretion and results in overly elongated tubes. Embryos lacking all three characterized claudins have tracheal phenotypes similar to any single mutant, indicating that these claudins act in the same pathway controlling tracheal tube length. However, we find that there are distinct requirements for these claudins in epithelial septate junction formation. Megatrachea is predominantly required for correct localization of septate junction components, while Sinuous is predominantly required for maintaining normal levels of septate junction proteins. Kune-kune is required for both localization and levels. Double- and triple-mutant combinations of Sinuous and Megatrachea with Kune-kune resemble the Kune-kune single mutant, suggesting that Kune-kune has a more central role in septate junction formation than either Sinuous or Megatrachea.
Figures






Similar articles
-
The claudin-like megatrachea is essential in septate junctions for the epithelial barrier function in Drosophila.Dev Cell. 2003 Oct;5(4):611-20. doi: 10.1016/s1534-5807(03)00275-2. Dev Cell. 2003. PMID: 14536062
-
Sinuous is a Drosophila claudin required for septate junction organization and epithelial tube size control.J Cell Biol. 2004 Jan 19;164(2):313-23. doi: 10.1083/jcb.200309134. J Cell Biol. 2004. PMID: 14734539 Free PMC article.
-
The claudin Megatrachea protein complex.J Biol Chem. 2012 Oct 26;287(44):36756-65. doi: 10.1074/jbc.M112.399410. Epub 2012 Aug 28. J Biol Chem. 2012. PMID: 22930751 Free PMC article.
-
Claudins in occluding junctions of humans and flies.Trends Cell Biol. 2006 Apr;16(4):181-8. doi: 10.1016/j.tcb.2006.02.006. Epub 2006 Mar 14. Trends Cell Biol. 2006. PMID: 16537104 Review.
-
A junctional problem of apical proportions: epithelial tube-size control by septate junctions in the Drosophila tracheal system.Curr Opin Cell Biol. 2004 Oct;16(5):493-9. doi: 10.1016/j.ceb.2004.07.008. Curr Opin Cell Biol. 2004. PMID: 15363798 Review.
Cited by
-
Sinuous Is a Claudin Required for Locust Molt in Locusta migratoria.Genes (Basel). 2024 Jun 27;15(7):850. doi: 10.3390/genes15070850. Genes (Basel). 2024. PMID: 39062629 Free PMC article.
-
Formation and remodelling of septate junctions in the epidermis of isopod Porcellioscaber during development.Zookeys. 2022 May 18;1101:159-181. doi: 10.3897/zookeys.1101.78711. eCollection 2022. Zookeys. 2022. PMID: 36760974 Free PMC article.
-
Crumbs organizes the transport machinery by regulating apical levels of PI(4,5)P2 in Drosophila.Elife. 2019 Nov 7;8:e50900. doi: 10.7554/eLife.50900. Elife. 2019. PMID: 31697234 Free PMC article.
-
Claudins reign: The claudin/EMP/PMP22/γ channel protein family in C. elegans.Tissue Barriers. 2013 Jul 1;1(3):e25502. doi: 10.4161/tisb.25502. Epub 2013 Jun 24. Tissue Barriers. 2013. PMID: 24665403 Free PMC article. Review.
-
A dynamic paracellular pathway serves diuresis in mosquito Malpighian tubules.Ann N Y Acad Sci. 2012 Jul;1258(1):166-76. doi: 10.1111/j.1749-6632.2012.06527.x. Ann N Y Acad Sci. 2012. PMID: 22731730 Free PMC article.
References
-
- Asano, A., K. Asano, H. Sasaki, M. Furuse and S. Tsukita, 2003. Claudins in Caenorhabditis elegans: their distribution and barrier function in the epithelium. Curr. Biol. 13 1042–1046. - PubMed
-
- Auld, V. J., R. D. Fetter, K. Broadie and C. S. Goodman, 1995. Gliotactin, a novel transmembrane protein on peripheral glia, is required to form the blood-nerve barrier in Drosophila. Cell 81 757–767. - PubMed
-
- Baumgartner, S., J. T. Littleton, K. Broadie, M. A. Bhat, R. Harbecke et al., 1996. A Drosophila neurexin is required for septate junction and blood-nerve barrier formation and function. Cell 87 1059–1068. - PubMed
-
- Behr, M., D. Riedel and R. Schuh, 2003. The claudin-like megatrachea is essential in septate junctions for the epithelial barrier function in Drosophila. Dev. Cell 5 611–620. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
