

LPCAT1 regulates surfactant phospholipid synthesis and is required for transitioning to air breathing in mice
- PMID: 20407208
- PMCID: PMC2860922
- DOI: 10.1172/JCI38061
LPCAT1 regulates surfactant phospholipid synthesis and is required for transitioning to air breathing in mice
Erratum in
- J Clin Invest. 2010 Jun 1;120(6):2248
Abstract
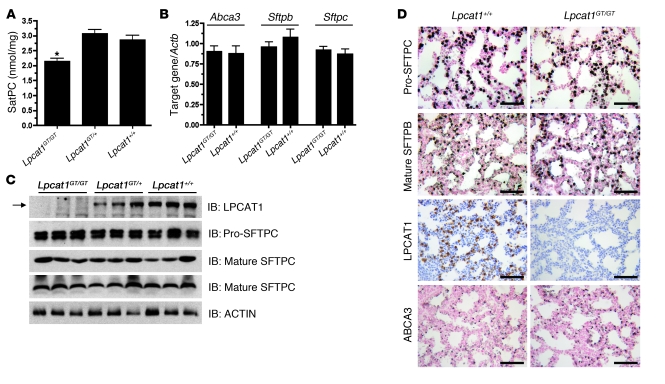
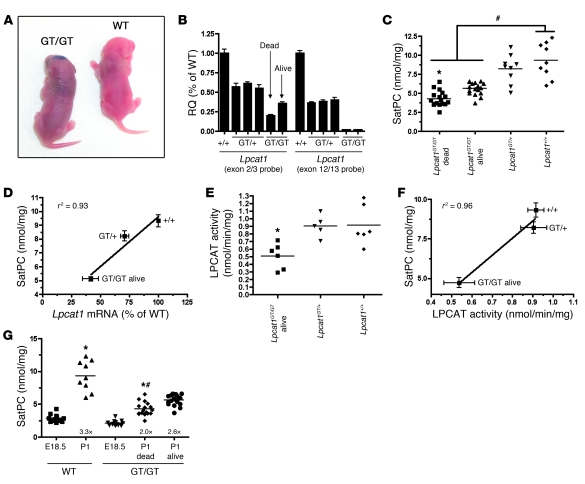
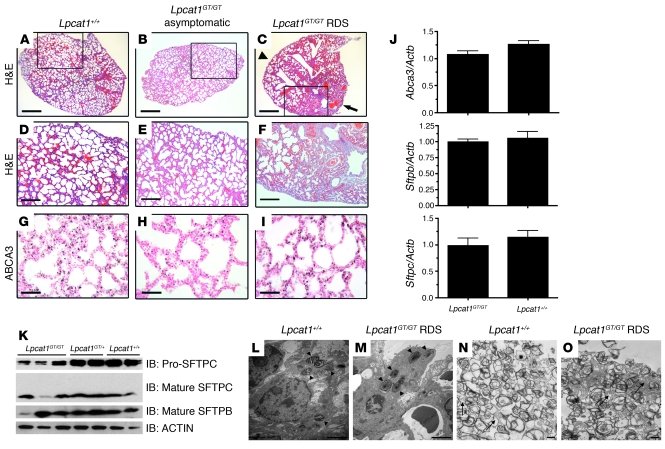
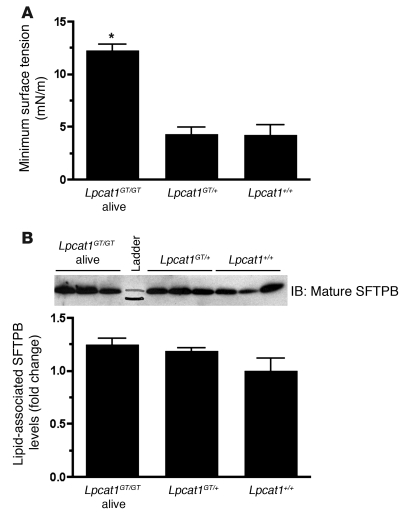
Respiratory distress syndrome (RDS), which is the leading cause of death in premature infants, is caused by surfactant deficiency. The most critical and abundant phospholipid in pulmonary surfactant is saturated phosphatidylcholine (SatPC), which is synthesized in alveolar type II cells de novo or by the deacylation-reacylation of existing phosphatidylcholine species. We recently cloned and partially characterized a mouse enzyme with characteristics of a lung lysophosphatidylcholine acyltransferase (LPCAT1) that we predicted would be involved in surfactant synthesis. Here, we describe our studies investigating whether LPCAT1 is required for pulmonary surfactant homeostasis. To address this issue, we generated mice bearing a hypomorphic allele of Lpcat1 (referred to herein as Lpcat1GT/GT mice) using a genetrap strategy. Newborn Lpcat1GT/GT mice showed varying perinatal mortality from respiratory failure, with affected animals demonstrating hallmarks of respiratory distress such as atelectasis and hyaline membranes. Lpcat1 mRNA levels were reduced in newborn Lpcat1GT/GT mice and directly correlated with SatPC content, LPCAT1 activity, and survival. Surfactant isolated from dead Lpcat1GT/GT mice failed to reduce minimum surface tension to wild-type levels. Collectively, these data demonstrate that full LPCAT1 activity is required to achieve the levels of SatPC essential for the transition to air breathing.
Figures
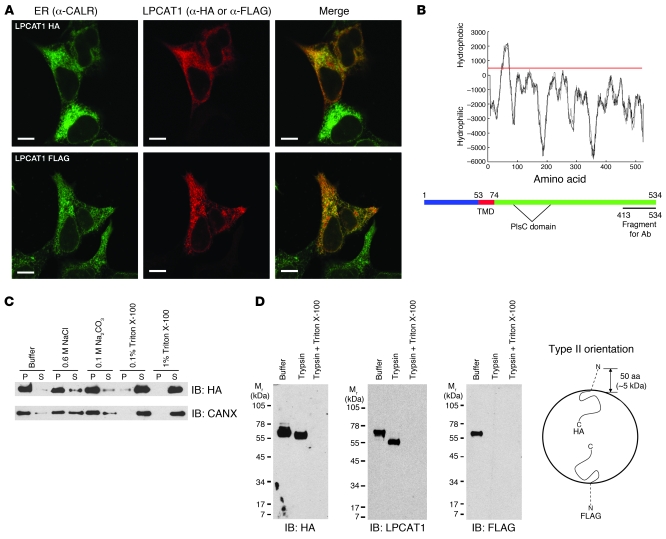
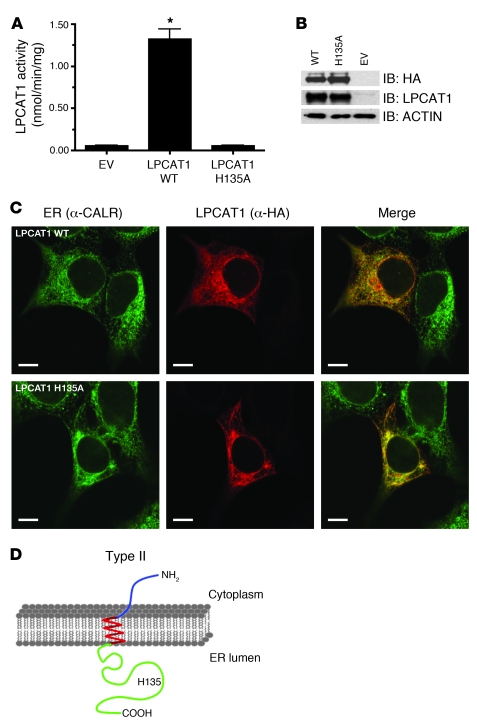
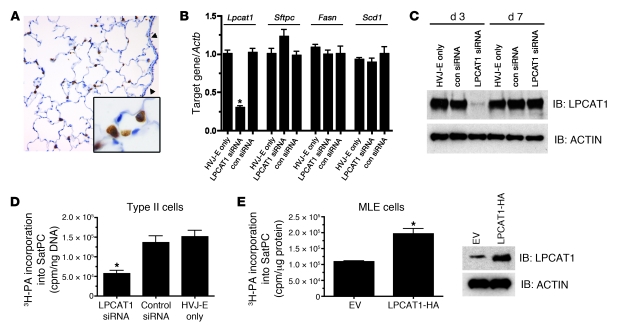
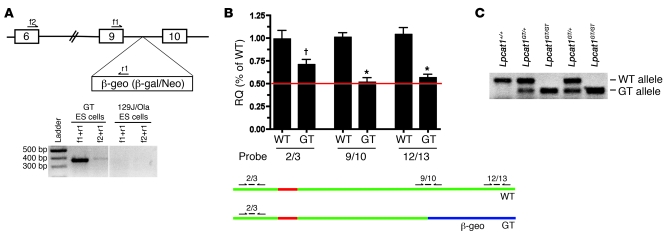
References
-
- Pison U, et al. Surfactant abnormalities in patients with respiratory failure after multiple trauma. Am Rev Respir Dis. 1989;140(4):1033–1039. - PubMed
-
- Gunther A, et al. Surfactant alterations in severe pneumonia, acute respiratory distress syndrome, and cardiogenic lung edema. Am J Respir Crit Care Med. 1996;153(1):176–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
