

TRF2/RAP1 and DNA-PK mediate a double protection against joining at telomeric ends
- PMID: 20407424
- PMCID: PMC2876953
- DOI: 10.1038/emboj.2010.49
TRF2/RAP1 and DNA-PK mediate a double protection against joining at telomeric ends
Abstract
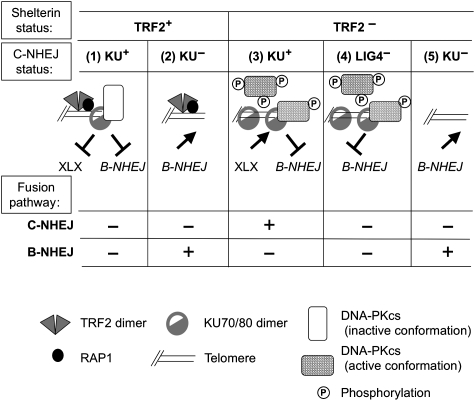
DNA-dependent protein kinase (DNA-PK) is a double-strand breaks repair complex, the subunits of which (KU and DNA-PKcs) are paradoxically present at mammalian telomeres. Telomere fusion has been reported in cells lacking these proteins, raising two questions: how is DNA-PK prevented from initiating classical ligase IV (LIG4)-dependent non-homologous end-joining (C-NHEJ) at telomeres and how is the backup end-joining (EJ) activity (B-NHEJ) that operates at telomeres under conditions of C-NHEJ deficiency controlled? To address these questions, we have investigated EJ using plasmid substrates bearing double-stranded telomeric tracks and human cell extracts with variable C-NHEJ or B-NHEJ activity. We found that (1) TRF2/RAP1 prevents C-NHEJ-mediated end fusion at the initial DNA-PK end binding and activation step and (2) DNA-PK counteracts a potent LIG4-independent EJ mechanism. Thus, telomeres are protected against EJ by a lock with two bolts. These results account for observations with mammalian models and underline the importance of alternative non-classical EJ pathways for telomere fusions in cells.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Both the classical and alternative non-homologous end joining pathways contribute to the fusion of drastically shortened telomeres induced by TRF2 overexpression.Cell Cycle. 2019 Apr;18(8):880-888. doi: 10.1080/15384101.2019.1598724. Epub 2019 Apr 6. Cell Cycle. 2019. PMID: 30907229 Free PMC article.
-
Human RAP1 inhibits non-homologous end joining at telomeres.EMBO J. 2009 Nov 4;28(21):3390-9. doi: 10.1038/emboj.2009.275. Epub 2009 Sep 17. EMBO J. 2009. PMID: 19763083 Free PMC article.
-
Chromosome end protection by RAP1-mediated inhibition of DNA-PK.Nature. 2025 Jun;642(8069):1090-1096. doi: 10.1038/s41586-025-08896-1. Epub 2025 Apr 16. Nature. 2025. PMID: 40240611 Free PMC article.
-
DNA-dependent protein kinase in telomere maintenance and protection.Cell Mol Biol Lett. 2020 Jan 17;25:2. doi: 10.1186/s11658-020-0199-0. eCollection 2020. Cell Mol Biol Lett. 2020. PMID: 31988640 Free PMC article. Review.
-
Mechanisms of DNA double strand break repair and chromosome aberration formation.Cytogenet Genome Res. 2004;104(1-4):14-20. doi: 10.1159/000077461. Cytogenet Genome Res. 2004. PMID: 15162010 Review.
Cited by
-
Unraveling secrets of telomeres: one molecule at a time.DNA Repair (Amst). 2014 Aug;20:142-153. doi: 10.1016/j.dnarep.2014.01.012. Epub 2014 Feb 22. DNA Repair (Amst). 2014. PMID: 24569170 Free PMC article. Review.
-
Depletion of Ku70/80 reduces the levels of extrachromosomal telomeric circles and inhibits proliferation of ALT cells.Aging (Albany NY). 2011 Apr;3(4):395-406. doi: 10.18632/aging.100308. Aging (Albany NY). 2011. PMID: 21512205 Free PMC article.
-
Trypanosoma brucei RAP1 Has Essential Functional Domains That Are Required for Different Protein Interactions.mSphere. 2020 Feb 26;5(1):e00027-20. doi: 10.1128/mSphere.00027-20. mSphere. 2020. PMID: 32102938 Free PMC article.
-
TRF2 interaction with Ku heterotetramerization interface gives insight into c-NHEJ prevention at human telomeres.Cell Rep. 2013 Oct 17;5(1):194-206. doi: 10.1016/j.celrep.2013.08.040. Epub 2013 Oct 3. Cell Rep. 2013. PMID: 24095731 Free PMC article.
-
A G-quadruplex structure within the 5'-UTR of TRF2 mRNA represses translation in human cells.Nucleic Acids Res. 2010 Nov;38(20):7187-98. doi: 10.1093/nar/gkq563. Epub 2010 Jun 22. Nucleic Acids Res. 2010. PMID: 20571083 Free PMC article.
References
-
- Amiard S, Doudeau M, Pinte S, Poulet A, Lenain C, Faivre-Moskalenko C, Angelov D, Hug N, Vindigni A, Bouvet P, Paoletti J, Gilson E, Giraud-Panis MJ (2007) A topological mechanism for TRF2-enhanced strand invasion. Nat Struct Mol Biol 14: 147–154 - PubMed
-
- Audebert M, Salles B, Calsou P (2004) Involvement of poly(ADP-ribose) polymerase-1 and XRCC1/DNA ligase III in an alternative route for DNA double-strand breaks rejoining. J Biol Chem 279: 55117–55126 - PubMed
-
- Bae NS, Baumann P (2007) A RAP1/TRF2 complex inhibits nonhomologous end-joining at human telomeric DNA ends. Mol Cell 26: 323–334 - PubMed
-
- Bailey SM, Brenneman MA, Halbrook J, Nickoloff JA, Ullrich RL, Goodwin EH (2004) The kinase activity of DNA-PK is required to protect mammalian telomeres. DNA Repair (Amst) 3: 225–233 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
