

Mechanisms of odor-tracking: multiple sensors for enhanced perception and behavior
- PMID: 20407585
- PMCID: PMC2854573
- DOI: 10.3389/fncel.2010.00006
Mechanisms of odor-tracking: multiple sensors for enhanced perception and behavior
Abstract
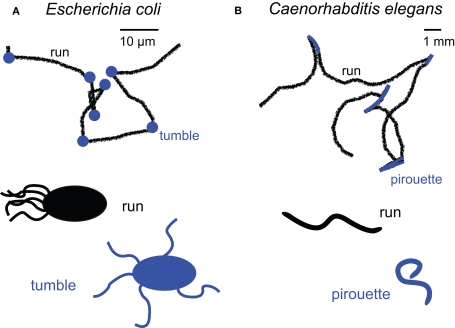
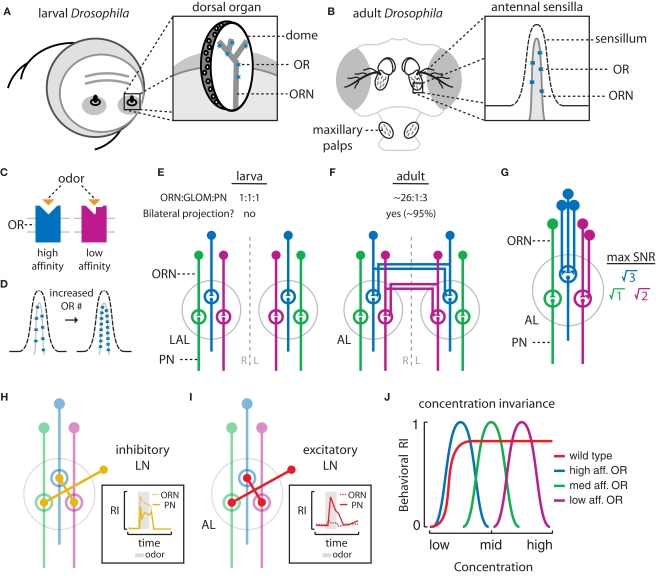
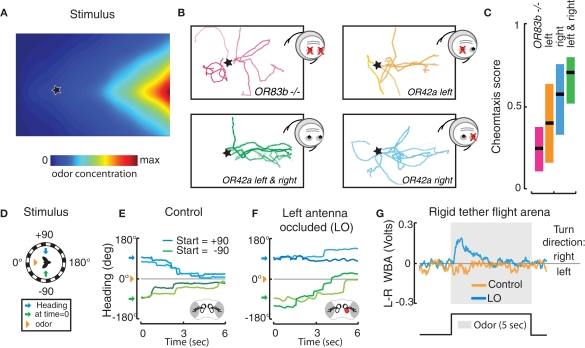
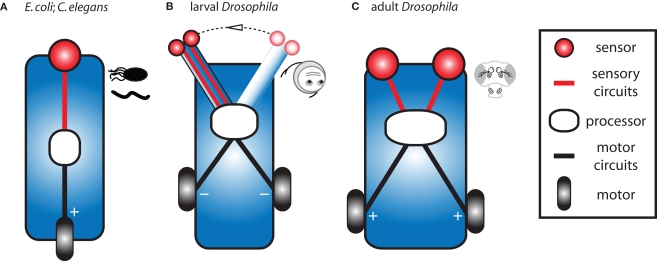
Early in evolution, the ability to sense and respond to changing environments must have provided a critical survival advantage to living organisms. From bacteria and worms to flies and vertebrates, sophisticated mechanisms have evolved to enhance odor detection and localization. Here, we review several modes of chemotaxis. We further consider the relevance of a striking and recurrent motif in the organization of invertebrate and vertebrate sensory systems, namely the existence of two symmetrical olfactory sensors. By combining our current knowledge about the olfactory circuits of larval and adult Drosophila, we examine the molecular and neural mechanisms underlying robust olfactory perception and extend these analyses to recent behavioral studies addressing the relevance and function of bilateral olfactory input for gradient detection. Finally, using a comparative theoretical approach based on Braitenberg's vehicles, we speculate about the relationships between anatomy, circuit architecture and stereotypical orientation behaviors.
Keywords: bilateral; chemotaxis; drosophila melanogaster; olfaction; orientation behavior; sensory perception.
Figures

References
-
- Arnold G., Masson C., Budharugsa S. (1985). Comparative study of the antennal lobes and their afferent pathway in the worker bee and the drone (Apis mellifera). Cell Tissue Res. 242, 593–605 10.1007/BF00225425 - DOI
LinkOut - more resources
Full Text Sources
Molecular Biology Databases