

Astrocytes impose postburst depression of release probability at hippocampal glutamate synapses
- PMID: 20410129
- PMCID: PMC6632337
- DOI: 10.1523/JNEUROSCI.3957-09.2010
Astrocytes impose postburst depression of release probability at hippocampal glutamate synapses
Abstract
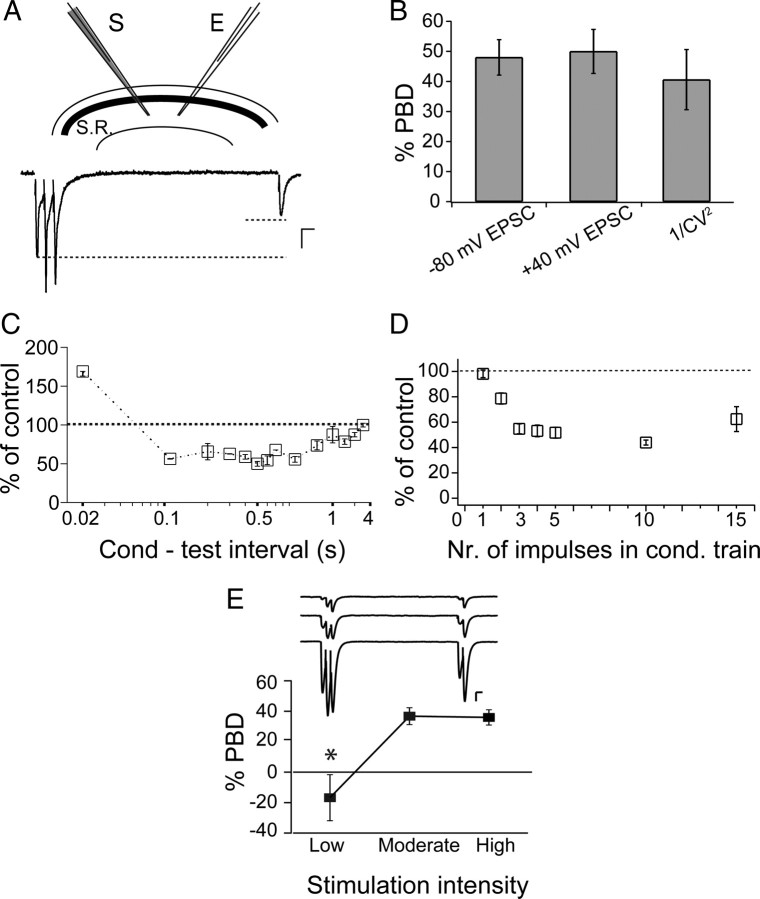
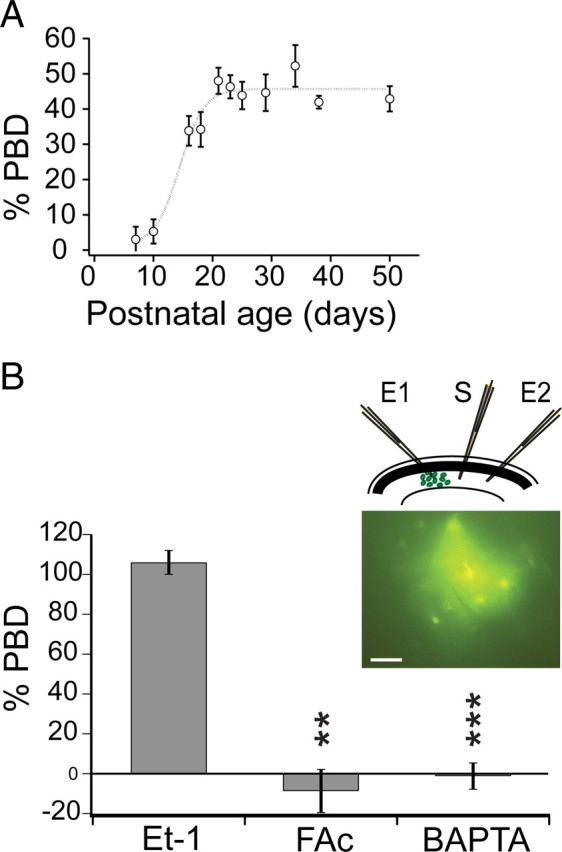
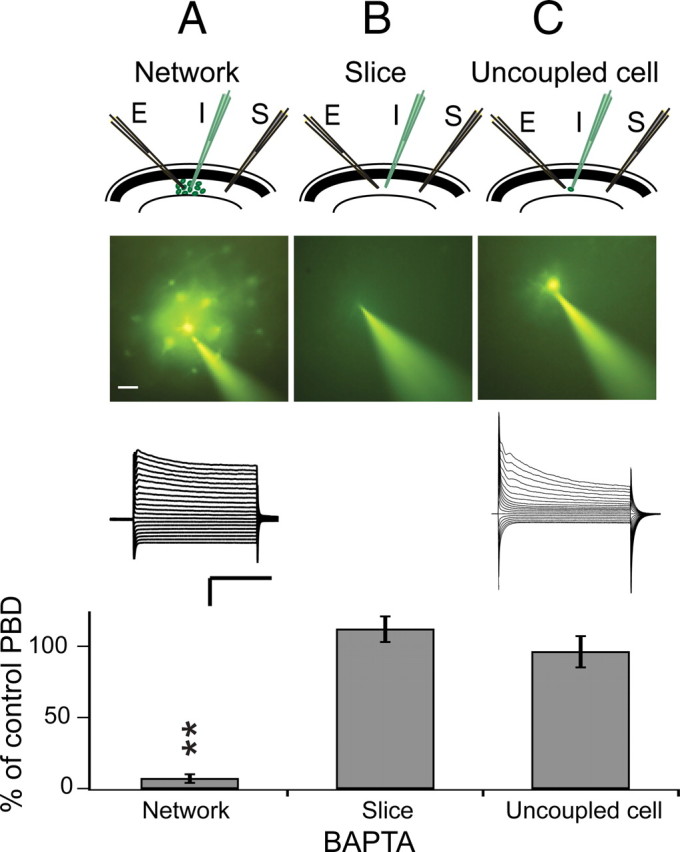
Many neurons typically fire action potentials in brief, high-frequency bursts with specific consequences for their synaptic output. Here we have examined short-term plasticity engaged during burst activation using electrophysiological recordings in acute rat hippocampal slices. We show that CA3-CA1 glutamate synapses exhibit a prominent depression of presynaptic release probability for approximately 1 s after such a burst. This postburst depression exhibits a distinct cooperativity threshold, is abolished by inhibiting astrocyte metabolism and astrocyte calcium signaling, and is not operational in the developing hippocampus. Our results suggest that astrocytes are actively involved in short-term synaptic depression, shaping synaptic activity during behaviorally relevant neural activity.
Figures
References
-
- Abbott LF, Regehr WG. Synaptic computation. Nature. 2004;431:796–803. - PubMed
-
- Andersen P. Interhippocampal impulses. II. Apical dendritic activation of CAI neurons. Acta Physiol Scand. 1960;48:178–208. - PubMed
-
- Araque A, Parpura V, Sanzgiri RP, Haydon PG. Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci. 1999;22:208–215. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous