

Presynaptic and postsynaptic mechanisms of synaptic plasticity and metaplasticity during intermediate-term memory formation in Aplysia
- PMID: 20410130
- PMCID: PMC6632334
- DOI: 10.1523/JNEUROSCI.4947-09.2010
Presynaptic and postsynaptic mechanisms of synaptic plasticity and metaplasticity during intermediate-term memory formation in Aplysia
Abstract
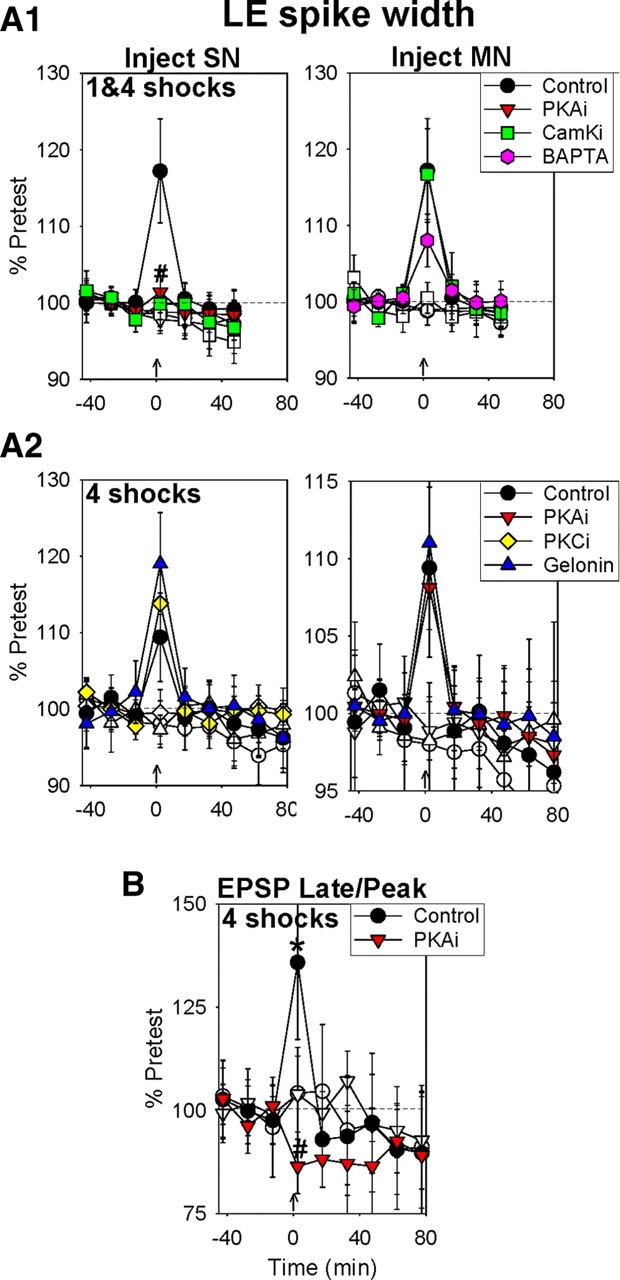
Synaptic plasticity and learning involve different mechanisms depending on the following: (1) the stage of plasticity and (2) the history of plasticity, or metaplasticity. However, little is known about how these two factors are related. We have addressed that question by examining mechanisms of synaptic plasticity during short-term and intermediate-term behavioral sensitization and dishabituation in a semi-intact preparation of the Aplysia siphon-withdrawal reflex. Dishabituation differs from sensitization in that it is preceded by habituation, and is thus a paradigm for metaplasticity. We find that whereas facilitation during short-term sensitization by one tail shock involves presynaptic covalent modifications by protein kinase A (PKA) and CamKII, facilitation during intermediate-term sensitization by four shocks involves both presynaptic (PKA, CaMKII) and postsynaptic (Ca(2+), CaMKII) covalent modifications, as well as both presynaptic and postsynaptic protein synthesis. The facilitation also involves presynaptic spike broadening 2.5 min after either one or four shocks, but not at later times. Dishabituation by four shocks differs from sensitization in several ways. First, it does not involve PKA or CaMKII, but rather involves presynaptic PKC. In addition, unlike sensitization with the same shock, dishabituation by four shocks does not involve protein synthesis or presynaptic spike broadening, and it also does not involve postsynaptic Ca(2+). These results demonstrate that not only the mechanisms but also the site of plasticity depend on both the stage of plasticity and metaplasticity during memory formation.
Figures





References
-
- Abraham WC, Bear MF. Metaplasticity: the plasticity of synaptic plasticity. Trends Neurosci. 1996;19:126–130. - PubMed
-
- Antonov I, Antonova I, Kandel ER, Hawkins RD. Activity-dependent presynaptic facilitation and Hebbian LTP are both required and interact during classical conditioning in Aplysia. Neuron. 2003;37:135–147. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous