

Heme binding to the Mammalian circadian clock protein period 2 is nonspecific
- PMID: 20411915
- PMCID: PMC2873066
- DOI: 10.1021/bi901945w
Heme binding to the Mammalian circadian clock protein period 2 is nonspecific
Abstract
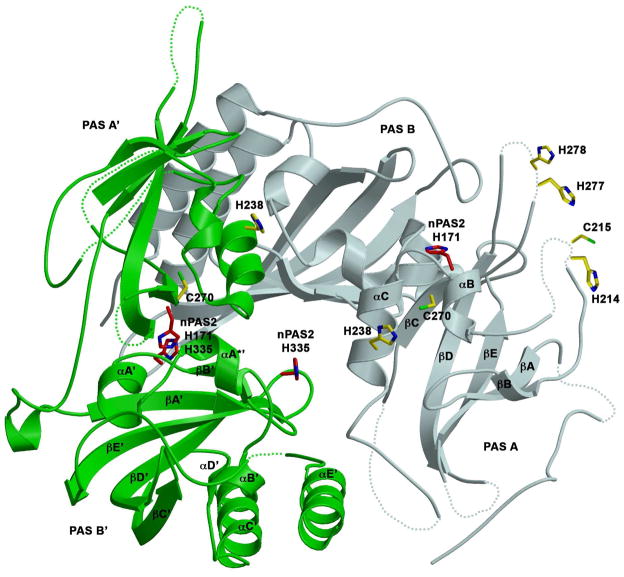
The mammalian circadian clock synchronizes physical and metabolic activity with the diurnal cycle through a transcriptional-posttranslational feedback loop. An additional feedback mechanism regulating clock timing has been proposed to involve oscillation in heme availability. Period 2 (PER2), an integral component in the negative feedback loop that establishes circadian rhythms in mammals, has been identified as a heme-binding protein. However, the majority of evidence for heme binding is based upon in vitro heme-binding assays. We sought to ascertain if these largely spectral assays could distinguish between specific and nonspecific heme interactions. Heme-binding properties by a number of other well-characterized proteins, all with no known biological role involving heme interaction, corresponded to those displayed by PER2. Site-directed mutants of putative heme-binding residues identified by MCD were unable to locate a specific heme-binding site on PER2. Protein film electrochemistry also indicates that heme binds PER2 nonspecifically on the protein surface. Our results establish the inability of qualitative in vitro assays to easily distinguish between specific and nonspecific heme binding. We conclude that heme binding to PER2 is likely to be nonspecific and does not involve the hydrophobic pocket within the PER2 PAS domains that in other PAS proteins commonly recognizes cofactors. These findings also question the significance of in vivo studies that implicate heme interactions with the clock proteins PER2 and nPAS2 in biological function.
Figures
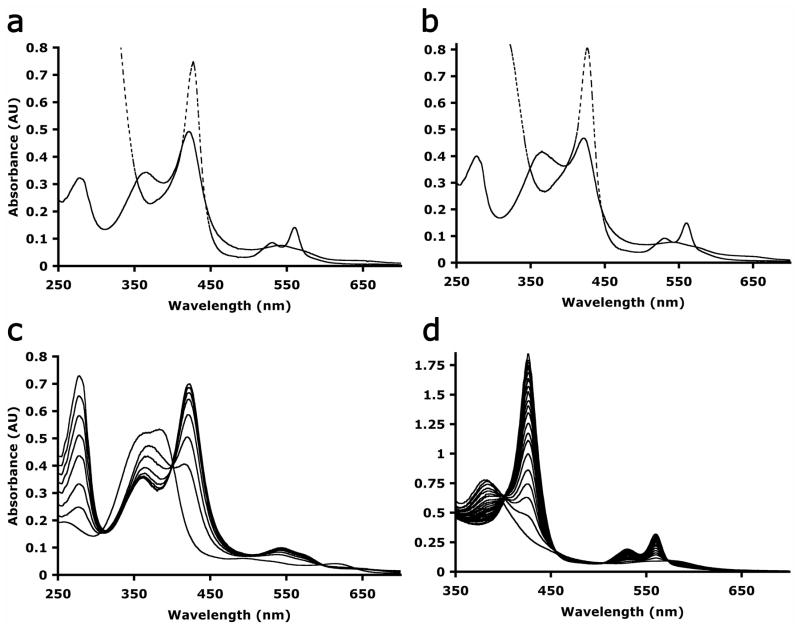
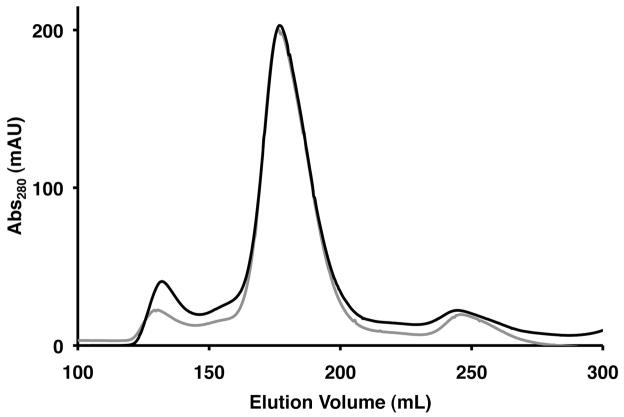
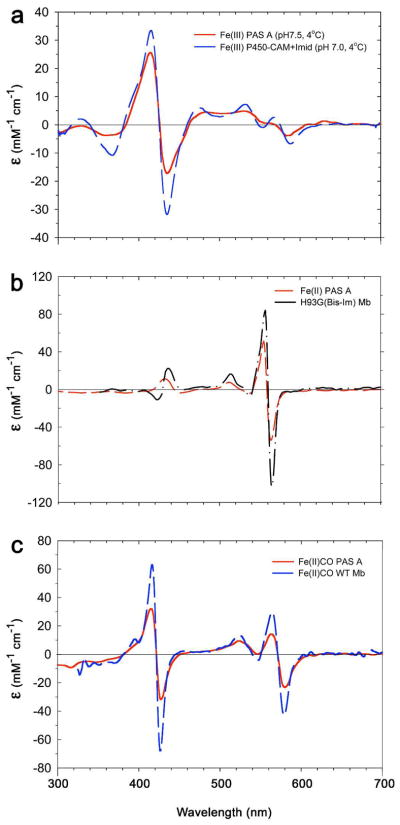

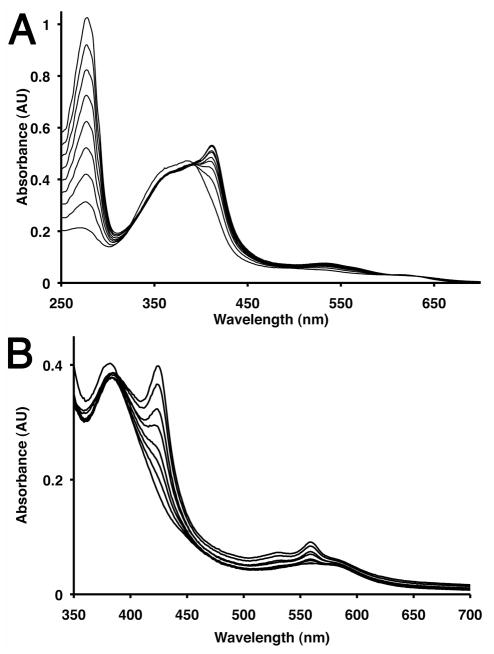
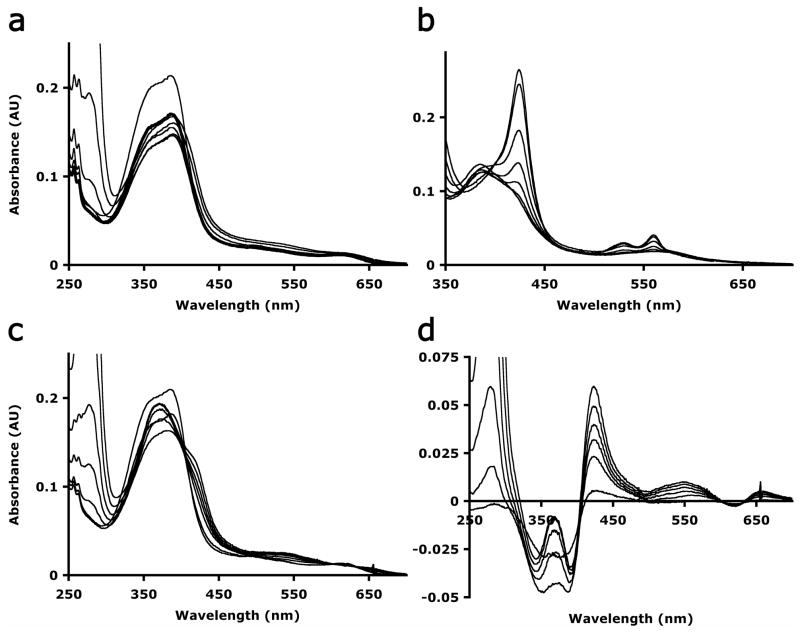
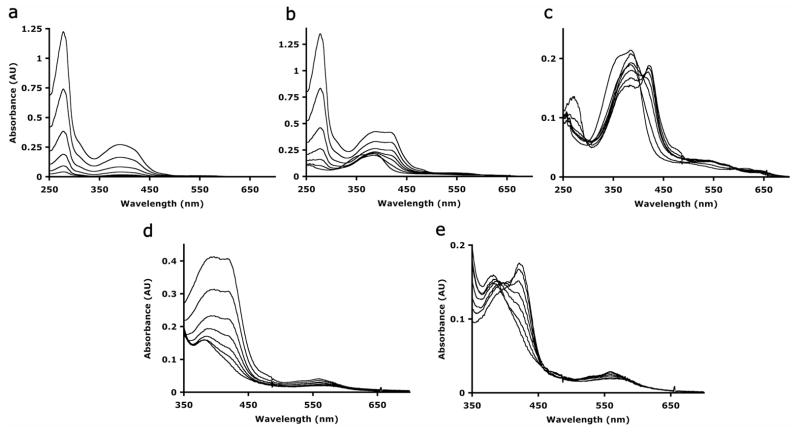
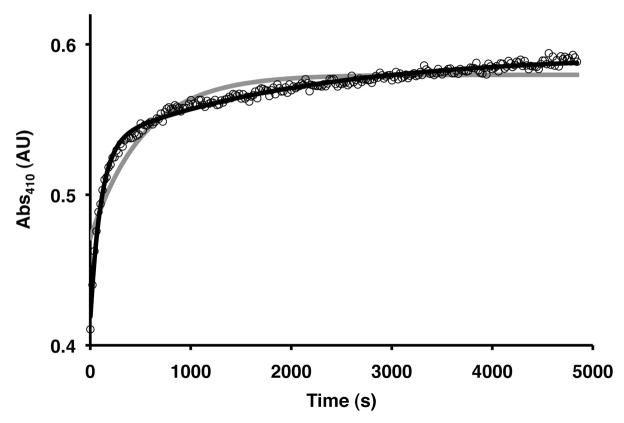
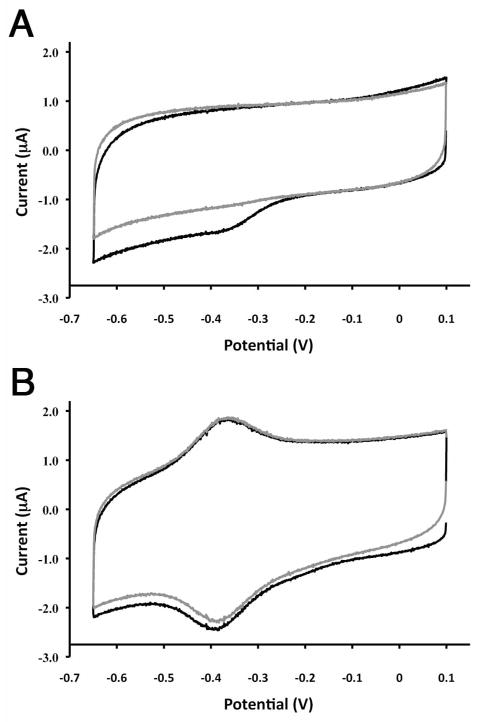
References
-
- Reppert SM, Weaver DR. Coordination of circadian timing in mammals. Nature. 2002;418:935–941. - PubMed
-
- Harms E, Kivimae S, Young MW, Saez L. Posttranscriptional and posttranslational regulation of clock genes. Journal of Biological Rhythms. 2004;19:361–373. - PubMed
-
- Young MW, Kay SA. Time zones: A comparative genetics of circadian clocks. Nature Reviews Genetics. 2001;2:702–715. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
