

Origin and fate of pseudogenes in Hemiascomycetes: a comparative analysis
- PMID: 20412590
- PMCID: PMC2876123
- DOI: 10.1186/1471-2164-11-260
Origin and fate of pseudogenes in Hemiascomycetes: a comparative analysis
Abstract
Background: Pseudogenes are ubiquitous genetic elements that derive from functional genes after mutational inactivation. Characterization of pseudogenes is important to understand genome dynamics and evolution, and its significance increases when several genomes of related organisms can be compared. Among yeasts, only the genome of the S. cerevisiae reference strain has been analyzed so far for pseudogenes.
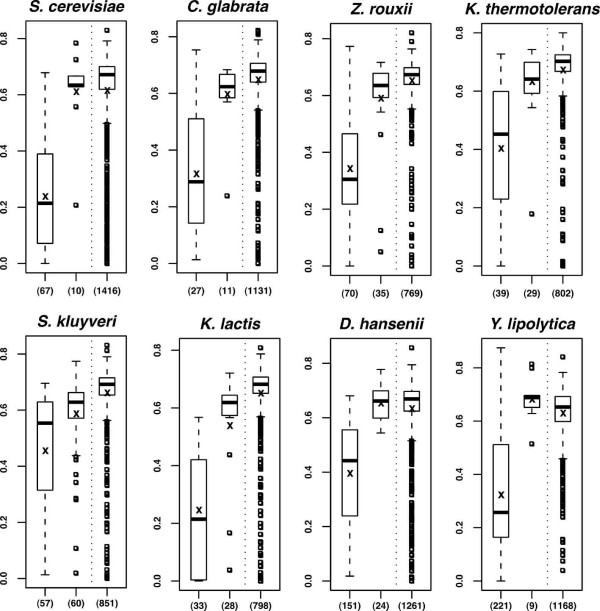
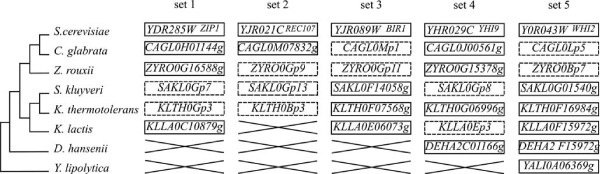
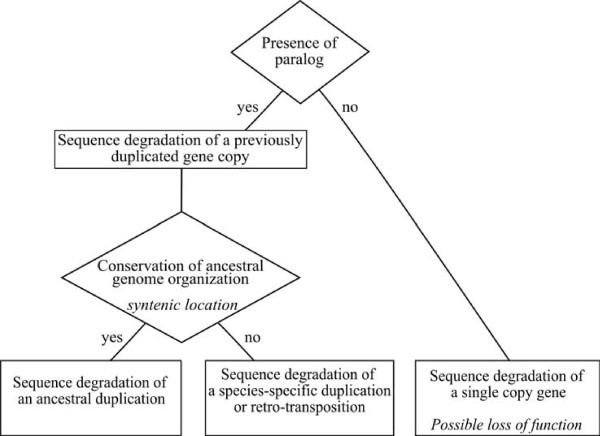
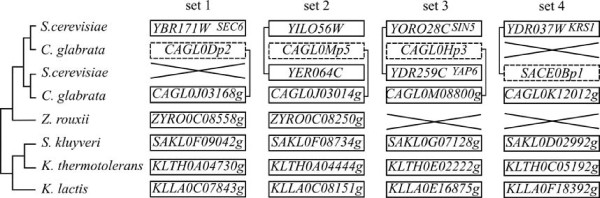
Results: We present here the first comparative analysis of pseudogenes within the fully sequenced and annotated genomes of eight yeast species, spanning the entire phylogenetic range of Hemiascomycetes. A total of 871 pseudogenes were found, out of which mutational degradation patterns and consequences on the genetic repertoire of each species could be identified. We found that most pseudogenes in yeasts originate from mutational degradation of gene copies formed after species-specific duplications but duplications of pseudogenes themselves are also encountered. In all yeasts, except in Y. lipolytica, pseudogenes tend to cluster in subtelomeric regions where they can outnumber the number of functional genes from 3 to 16 times. Pseudogenes are generally not conserved between the yeast species studied (except in two cases), consistent with their large evolutionary distances, but tend to be conserved among S. cerevisiae strains. Reiterated pseudogenization of some genes is often observed in different lineages and may affect functions essential in S. cerevisiae, which are, therefore, lost in other species. Although a variety of functions are affected by pseudogenization, there is a bias towards functions involved in the adaptation of the yeasts to their environment, and towards genes of unknown functions.
Conclusions: Our work illustrates for the first time the formation of pseudogenes in different branches of hemiascomycetous yeasts, showing their limited conservation and how they testify for the adaptation of the yeasts functional repertoires.
Figures

References
-
- Dasilva C, Hadji H, Ozouf-Costaz C, Nicaud S, Jaillon O, Weissenbach J, Crollius HR. Remarkable compartmentalization of transposable elements and pseudogenes in the heterochromatin of the Tetraodon nigroviridis genome. Proc Natl Acad Sci USA. 2002;99:13636–13641. doi: 10.1073/pnas.202284199. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
