

beta-catenin activity in the dermal papilla regulates morphogenesis and regeneration of hair
- PMID: 20412777
- PMCID: PMC2893731
- DOI: 10.1016/j.devcel.2010.01.016
beta-catenin activity in the dermal papilla regulates morphogenesis and regeneration of hair
Abstract
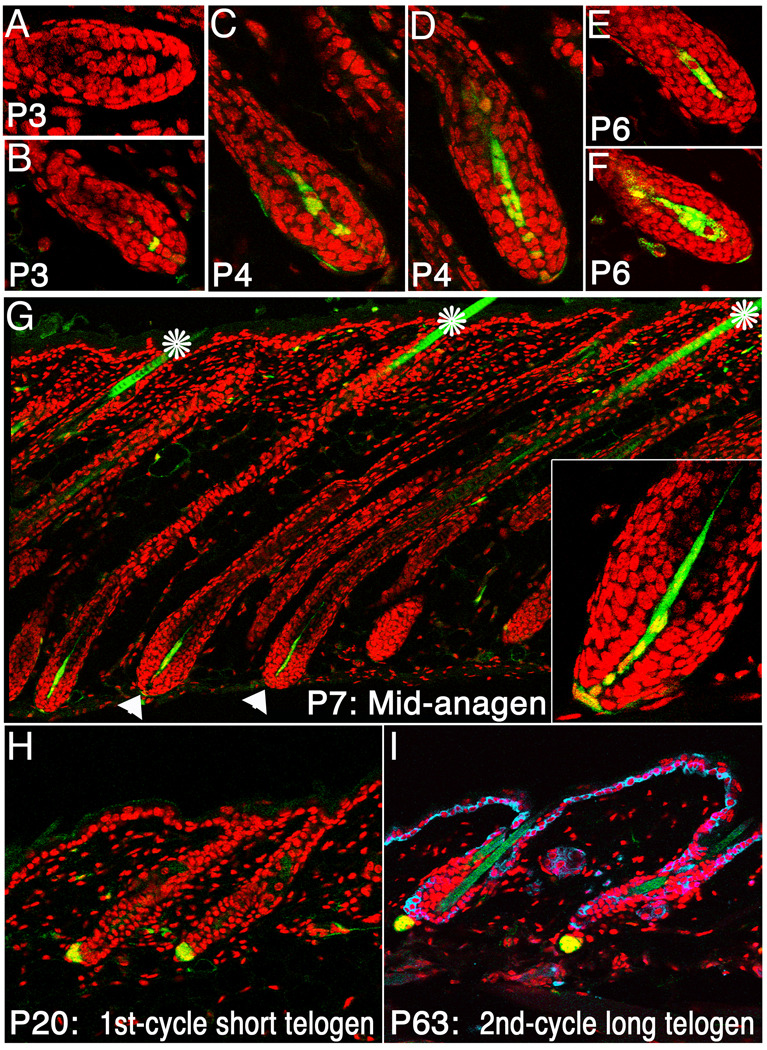
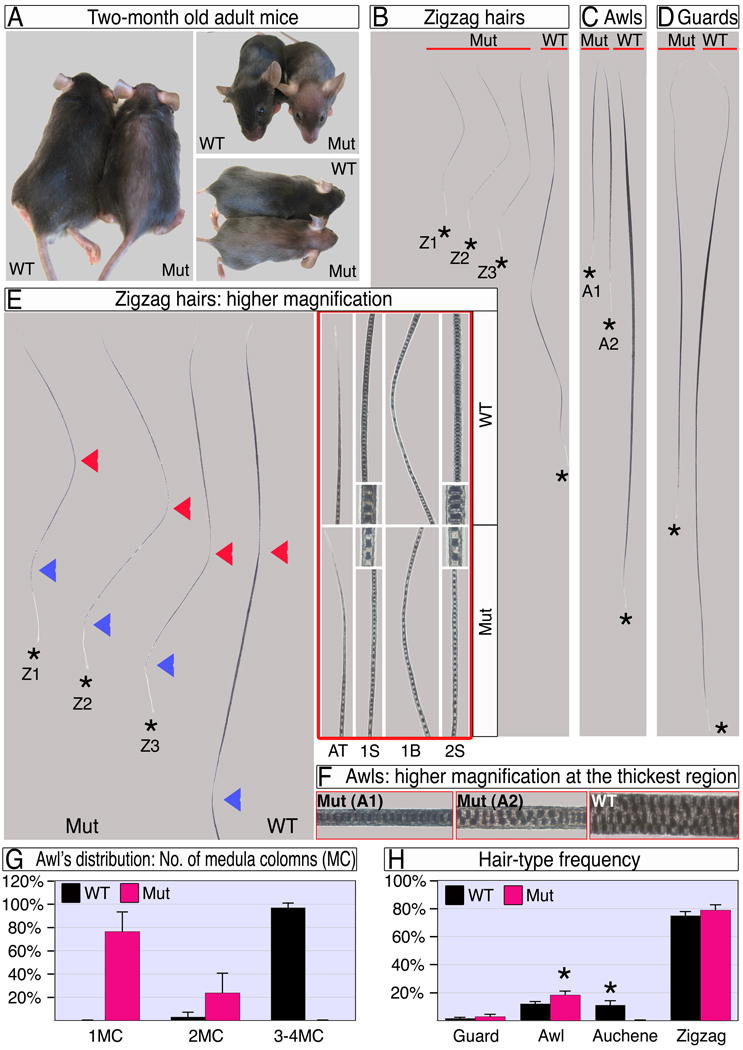
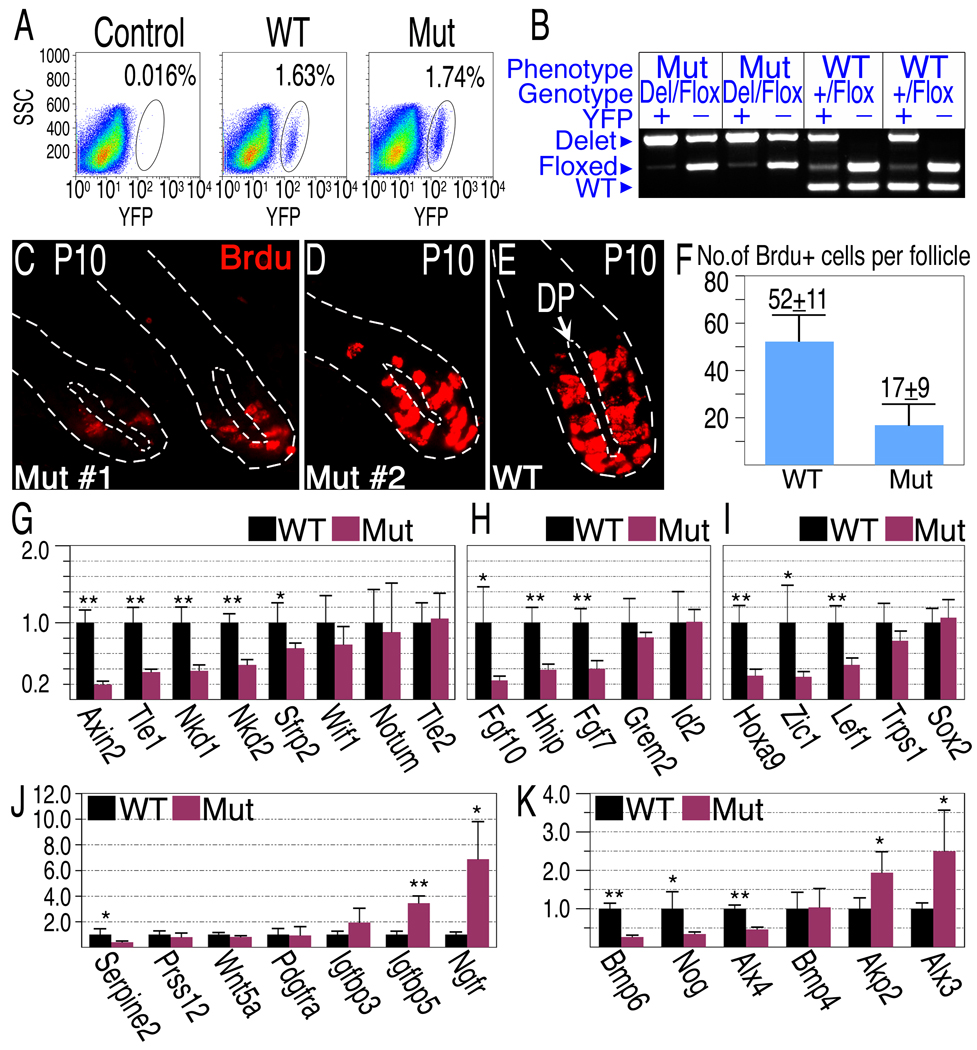
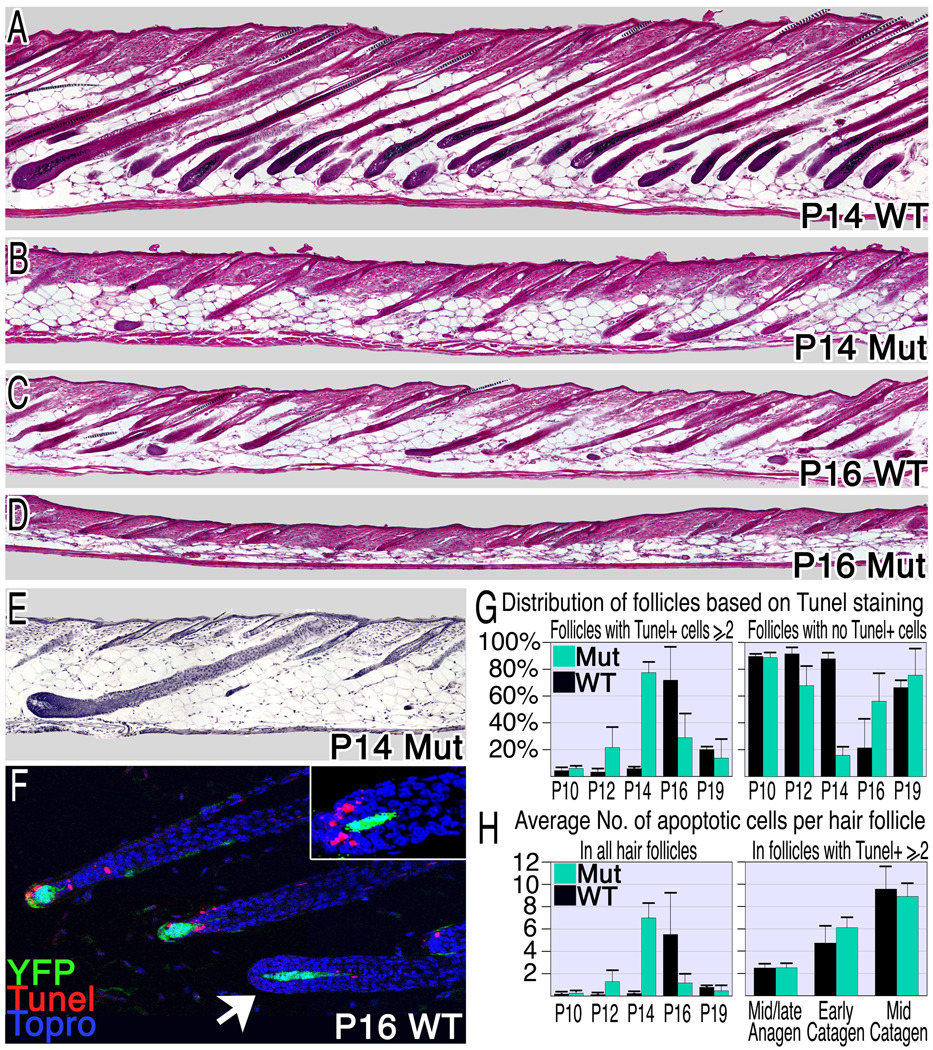
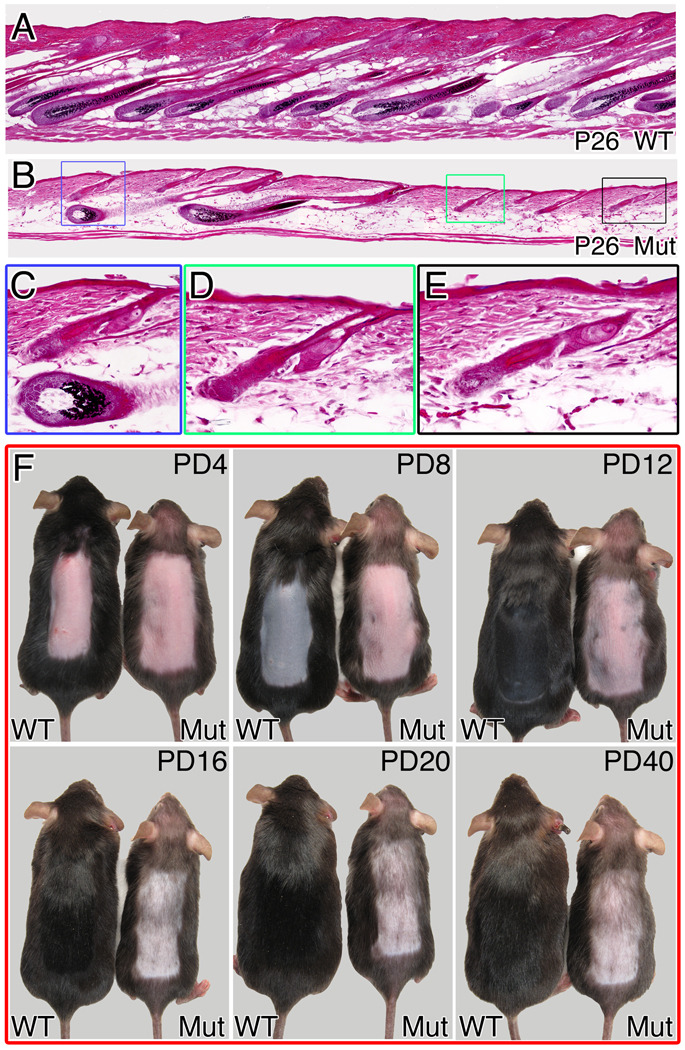
The activity of keratinocytes in the hair follicle is regulated by signals from a specialized mesenchymal niche, the dermal papilla (DP). Here, mice expressing cre recombinase in the DP were developed to probe the interaction between follicular keratinocytes and the DP in vivo. Inactivation of the beta-catenin gene within DP of fully developed hair follicles results in dramatically reduced proliferation of the progenitors and their progeny that generate the hair shaft, and, subsequently, premature induction of the destructive phase of the hair cycle. It also prevents regeneration of the cycling follicle from stem cells. Gene expression analysis reveals that beta-catenin activity in the DP regulates signaling pathways, including FGF and IGF, that can mediate the DP's inductive effects. This study reveals a signaling loop that employs Wnt/beta-catenin signaling in both epithelial progenitor cells and their mesenchymal niche to govern and coordinate the interactions between these compartments to guide hair morphogenesis.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Andl T, Reddy ST, Gaddapara T, Millar SE. WNT signals are required for the initiation of hair follicle development. Dev Cell. 2002;2:643–653. - PubMed
-
- Barreca A, De Luca M, Del Monte P, Bondanza S, Damonte G, Cariola G, Di Marco E, Giordano G, Cancedda R, Minuto F. In vitro paracrine regulation of human keratinocyte growth by fibroblast-derived insulin-like growth factors. J Cell Physiol. 1992;151:262–268. - PubMed
-
- Botchkarev VA, Botchkareva NV, Albers KM, Chen LH, Welker P, Paus R. A role for p75 neurotrophin receptor in the control of apoptosis-driven hair follicle regression. FASEB J. 2000;14:1931–1942. - PubMed
-
- Brault V, Moore R, Kutsch S, Ishibashi M, Rowitch DH, McMahon AP, Sommer L, Boussadia O, Kemler R. Inactivation of the beta-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development. 2001;128:1253–1264. - PubMed
-
- DasGupta R, Fuchs E. Multiple roles for activated LEF/TCF transcription complexes during hair follicle development and differentiation. Development. 1999;126:4557–4568. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
