

An endogenous, systemic RNAi pathway in plants
- PMID: 20414198
- PMCID: PMC2876969
- DOI: 10.1038/emboj.2010.65
An endogenous, systemic RNAi pathway in plants
Retraction in
-
Retraction: 'An endogenous, systemic RNAi pathway in plants'.EMBO J. 2015 Oct 14;34(20):2596. doi: 10.15252/embj.201570040. Epub 2015 Aug 19. EMBO J. 2015. PMID: 26290336 Free PMC article. No abstract available.
Abstract
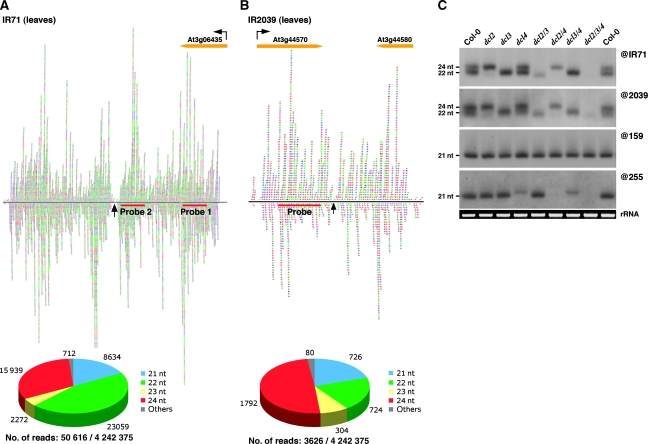
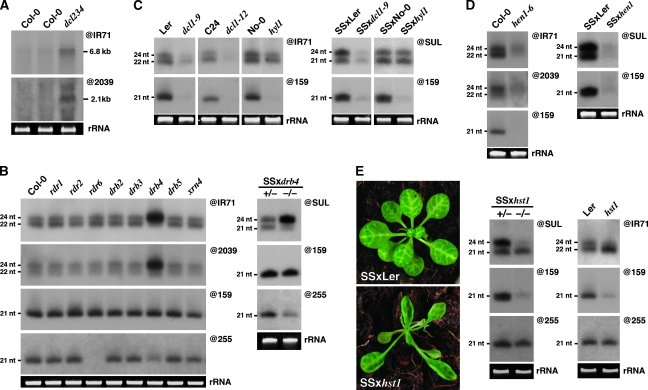
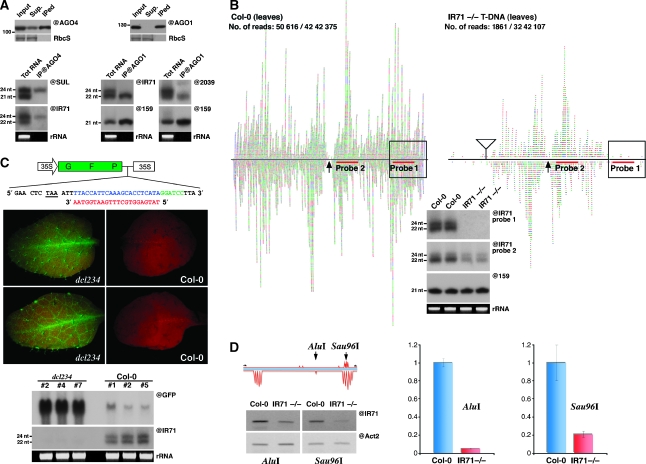
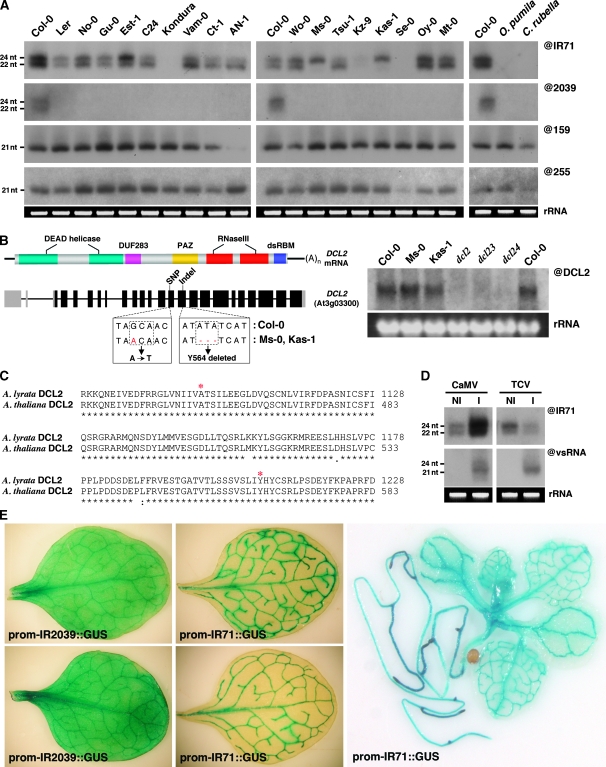
Recent work on metazoans has uncovered the existence of an endogenous RNA-silencing pathway that functionally recapitulates the effects of experimental RNA interference (RNAi) used for gene knockdown in organisms such as Caenorhabditis elegans and Drosophila. The endogenous short interfering (si)RNA involved in this pathway are processed by Dicer-like nucleases from genomic loci re-arranged to form extended inverted repeats (IRs) that produce perfect or near-perfect dsRNA molecules. Although such IR loci are commonly detected in plant genomes, their genetics, evolution and potential contribution to plant biology through endogenous silencing have remained largely unexplored. Through an exhaustive analysis performed using Arabidopsis, we provide here evidence that at least two such endogenous IRs are genetically virtually indistinguishable from the transgene constructs commonly used for RNAi in plants. We show how these loci can be useful probes of the cellular mechanism and fluidity of RNA-silencing pathways in plants, and provide evidence that they may arise and disappear on an ecotype scale, show highly cell-specific expression patterns and respond to various stresses. IR loci thus have the potential to act as molecular sensors of the local environments found within distinct ecological plant niches. We further show that the various siRNA size classes produced by at least one of these IR loci are functionally loaded into cognate effector proteins and mediate both post-transcriptional gene silencing and RNA-directed DNA methylation (RdDM) of endogenous as well as exogenous targets. Finally, and as previously reported during plant experimental RNAi, we provide evidence that endogenous IR-derived siRNAs of all size classes are not cell-autonomous and can be transported through graft junctions over long distances, in target tissues where they are functional, at least in mediating RdDM. Collectively, these results define the existence of a bona fide, endogenous and systemic RNAi pathway in plants that may have implications in adaptation, epiallelism and trans-generational memory.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Adenot X, Elmayan T, Lauressergues D, Boutet S, Bouche N, Gasciolli V, Vaucheret H (2006) DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Curr Biol 16: 927–932 - PubMed
-
- Allen E, Xie Z, Gustafson AM, Sung GH, Spatafora JW, Carrington JC (2004) Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana. Nat Genet 36: 1282–1290 - PubMed
-
- Banks JA, Masson P, Fedoroff N (1988) Molecular mechanisms in the developmental regulation of the maize Suppressor-mutator transposable element. Genes Dev 2: 1364–1380 - PubMed
-
- Bartel DP (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281–297 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
