

Mutation of Asn28 disrupts the dimerization and enzymatic activity of SARS 3CL(pro)
- PMID: 20420403
- PMCID: PMC2877132
- DOI: 10.1021/bi1002585
Mutation of Asn28 disrupts the dimerization and enzymatic activity of SARS 3CL(pro)
Abstract
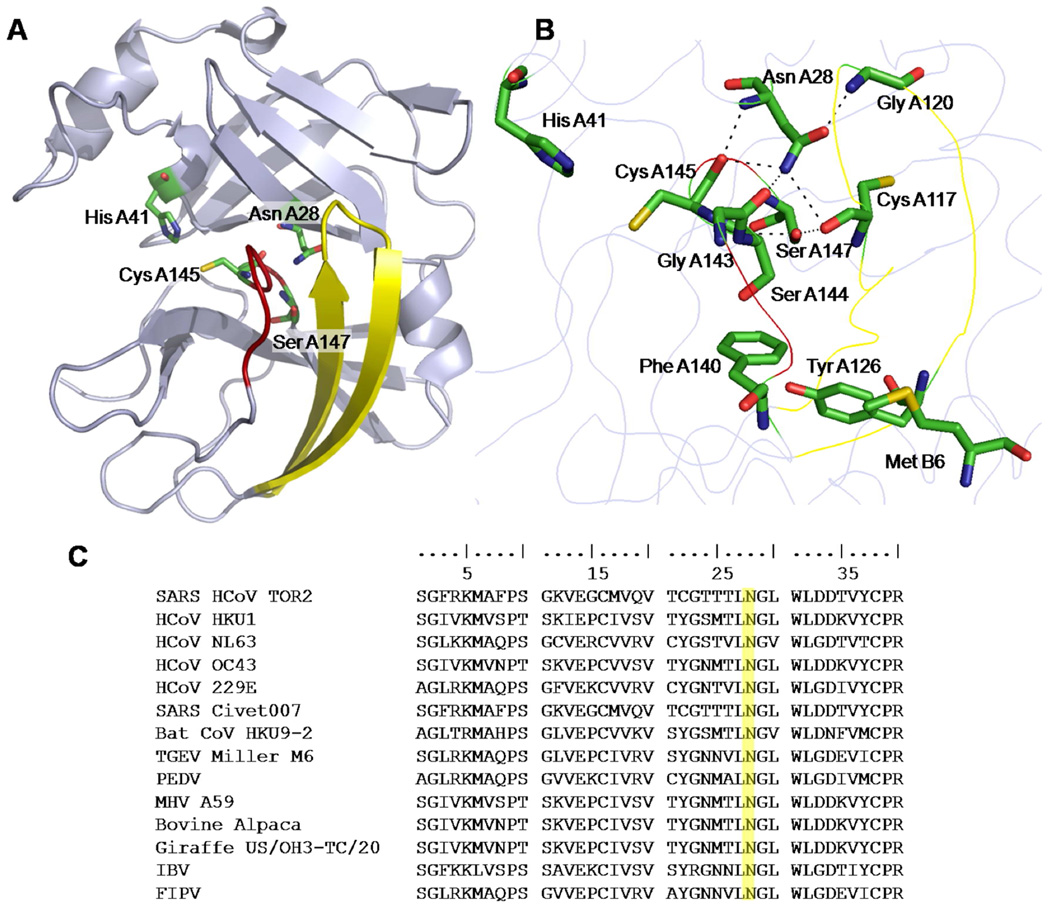
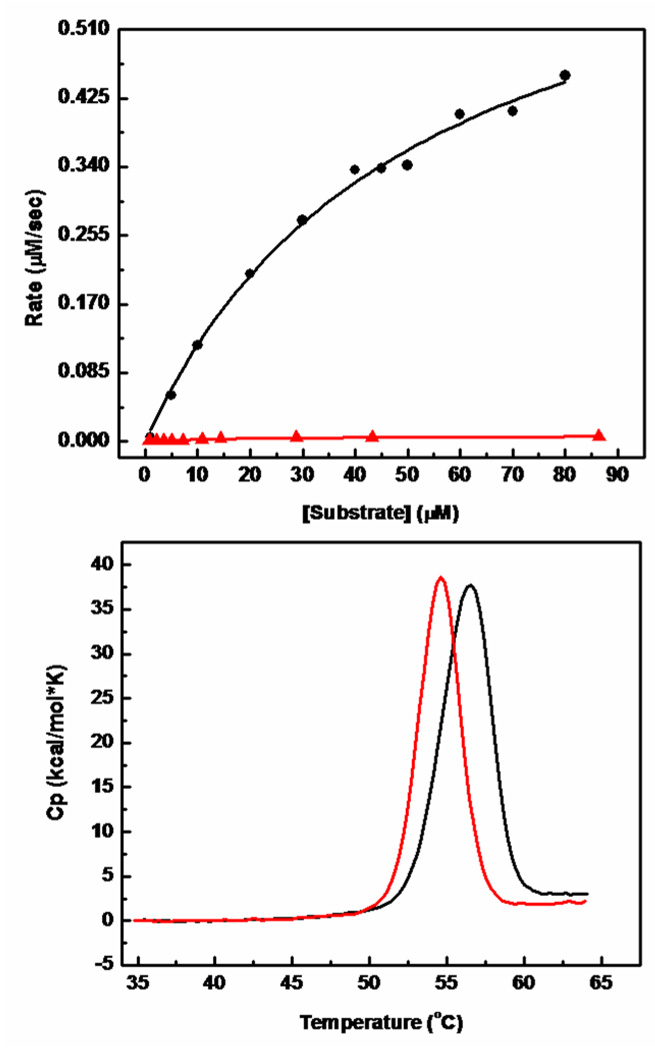
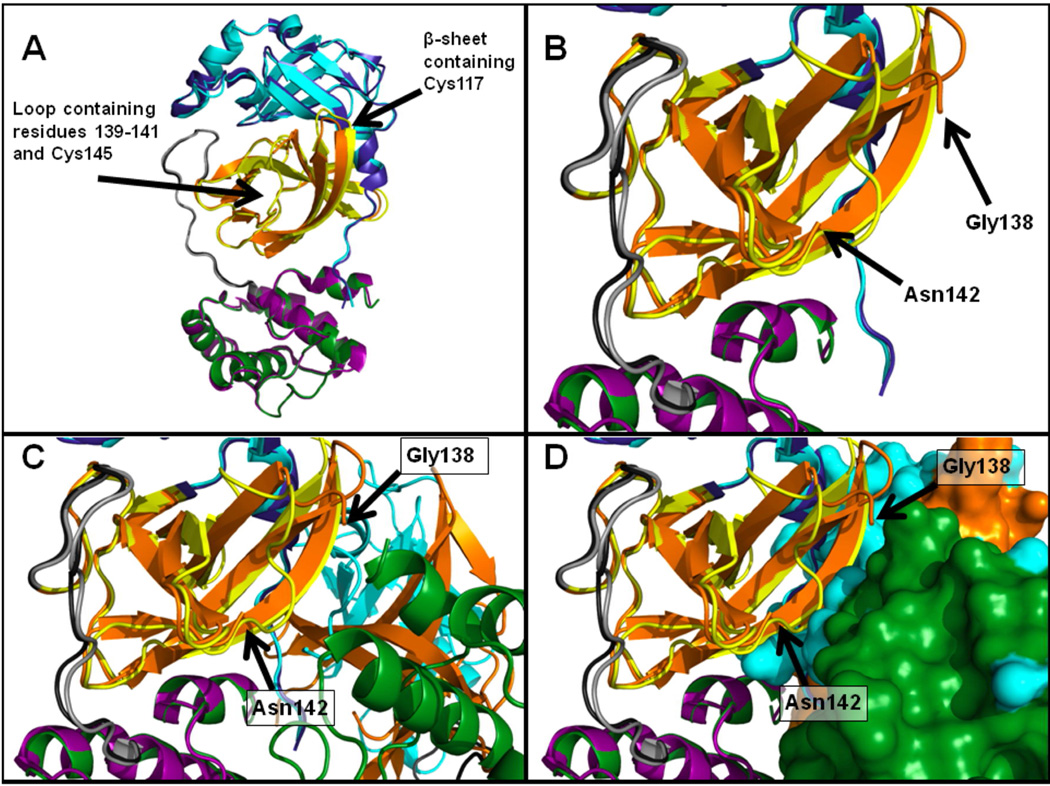
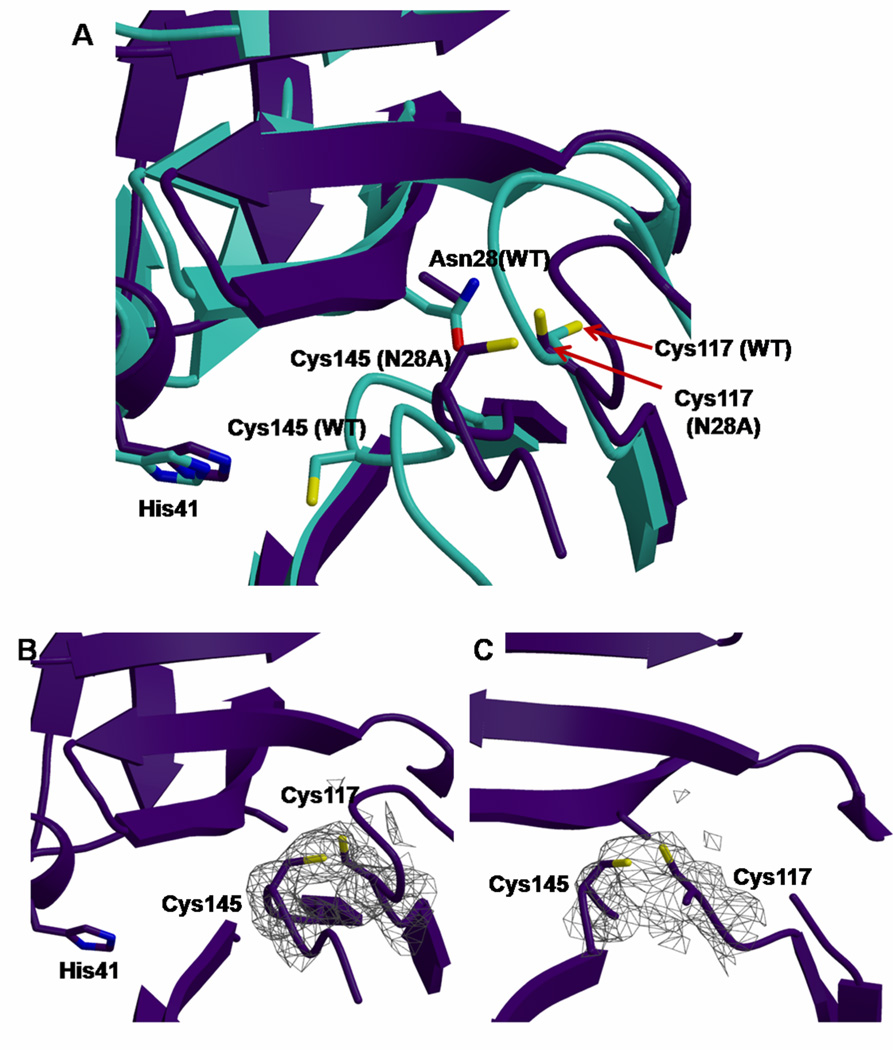
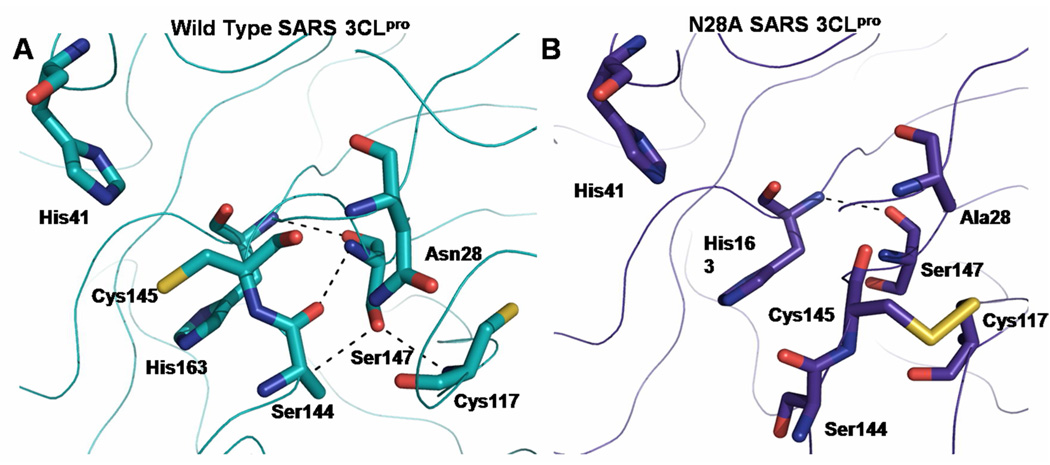
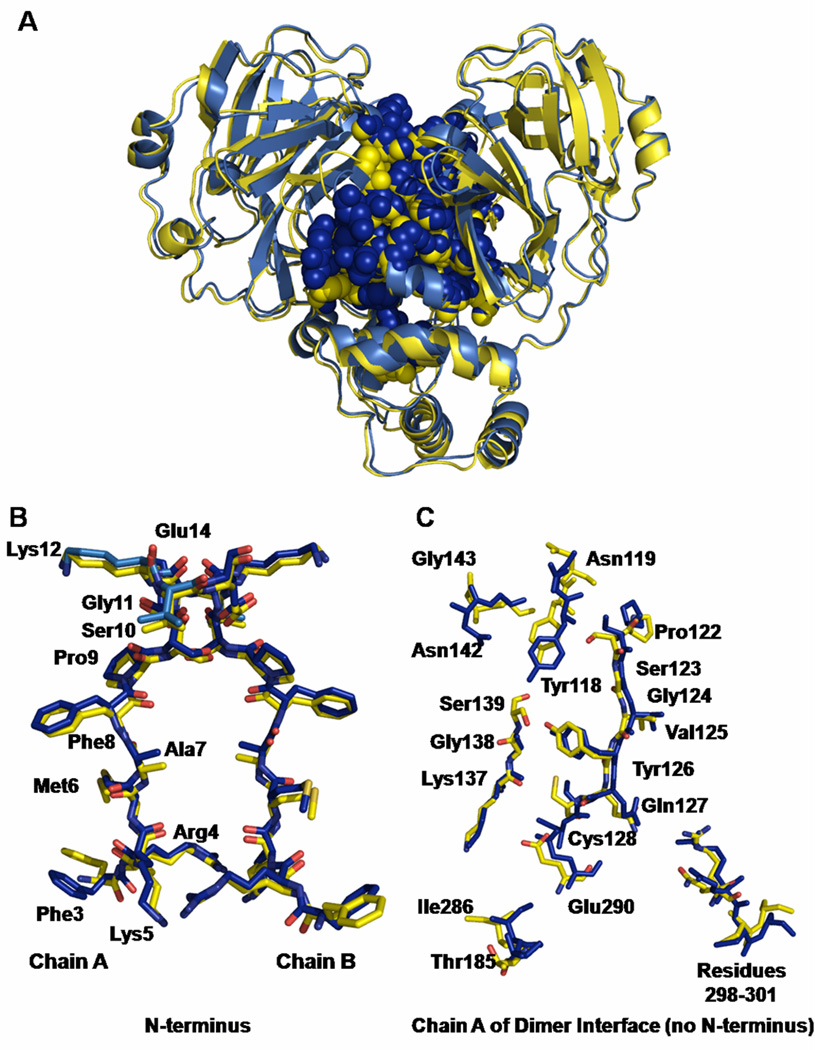
Coronaviruses are responsible for a significant proportion of annual respiratory and enteric infections in humans and other mammals. The most prominent of these viruses is the severe acute respiratory syndrome coronavirus (SARS-CoV) which causes acute respiratory and gastrointestinal infection in humans. The coronavirus main protease, 3CL(pro), is a key target for broad-spectrum antiviral development because of its critical role in viral maturation and high degree of structural conservation among coronaviruses. Dimerization is an indispensable requirement for the function of SARS 3CL(pro) and is regulated through mechanisms involving both direct and long-range interactions in the enzyme. While many of the binding interactions at the dimerization interface have been extensively studied, those that are important for long-range control are not well-understood. Characterization of these dimerization mechanisms is important for the structure-based design of new treatments targeting coronavirus-based infections. Here we report that Asn28, a residue 11 A from the closest residue in the opposing monomer, is essential for the enzymatic activity and dimerization of SARS 3CL(pro). Mutation of this residue to alanine almost completely inactivates the enzyme and results in a 19.2-fold decrease in the dimerization K(d). The crystallographic structure of the N28A mutant determined at 2.35 A resolution reveals the critical role of Asn28 in maintaining the structural integrity of the active site and in orienting key residues involved in binding at the dimer interface and substrate catalysis. These findings provide deeper insight into complex mechanisms regulating the activity and dimerization of SARS 3CL(pro).
Figures

References
-
- Drosten C, Gunther S, Preiser W, van der Werf S, Brodt H-R, Becker S, Rabenau H, Panning M, Kolesnikova L, Fouchier RAM, Berger A, Burguiere A-M, Cinatl J, Eickmann M, Escriou N, Grywna K, Kramme S, Manuguerra J-C, Muller S, Rickerts V, Sturmer M, Vieth S, Klenk H-D, Osterhaus ADME, Schmitz H, Doerr HW. Identification of a Novel Coronavirus in Patients with Severe Acute Respiratory Syndrome. N. Engl. J. Med. 2003;348:1967–1976. - PubMed
-
- Guan Y, Zheng BJ, He YQ, Liu XL, Zhuang ZX, Cheung CL, Luo SW, Li PH, Zhang LJ, Guan YJ, Butt KM, Wong KL, Chan KW, Lim W, Shortridge KF, Yuen KY, Peiris JSM, Poon LLM. Isolation and Characterization of Viruses Related to the SARS Coronavirus from Animals in Southern China. Science. 2003;302:276–278. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
