

Heterogeneity and hierarchy within the most primitive hematopoietic stem cell compartment
- PMID: 20421392
- PMCID: PMC2882827
- DOI: 10.1084/jem.20091318
Heterogeneity and hierarchy within the most primitive hematopoietic stem cell compartment
Abstract
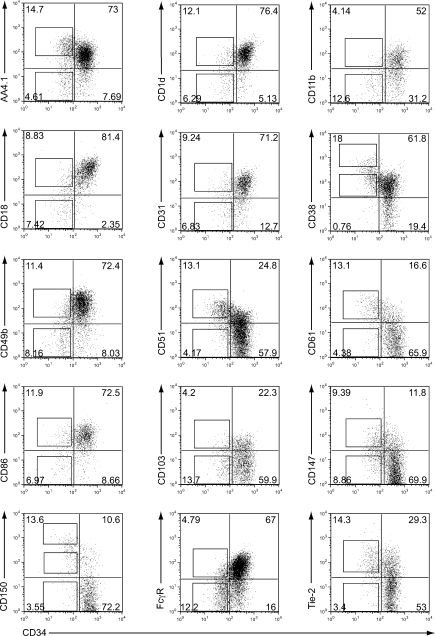
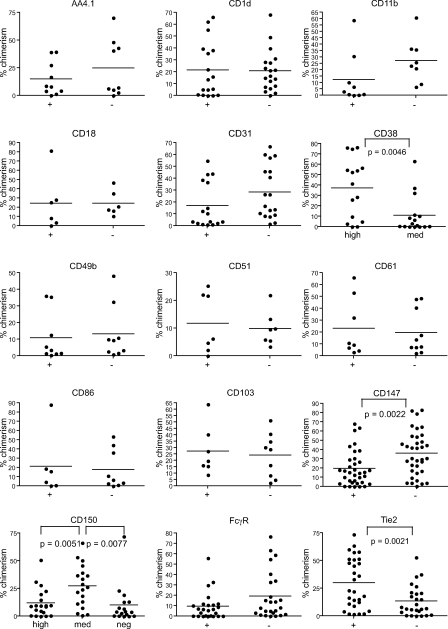
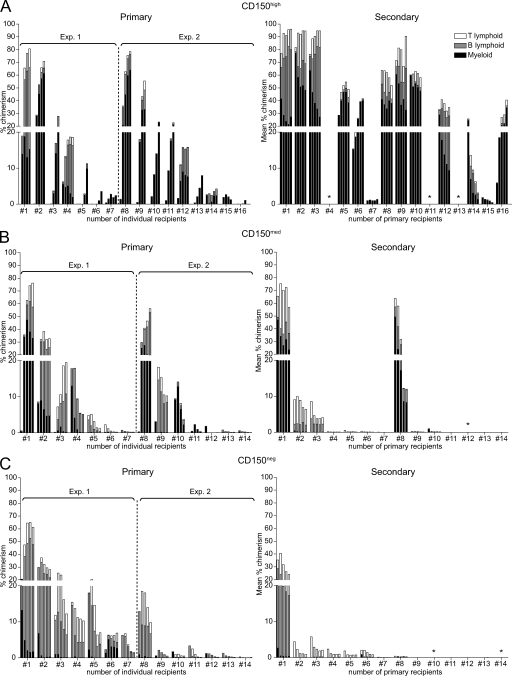
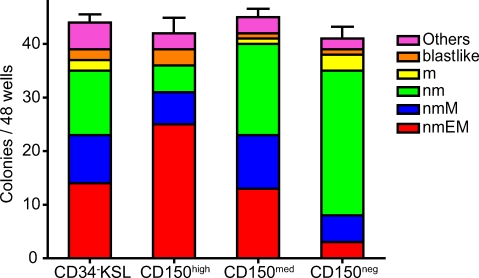
Hematopoietic stem cells (HSCs) have been extensively characterized based on functional definitions determined by experimental transplantation into lethally irradiated mice. In mice, HSCs are heterogeneous with regard to self-renewal potential, in vitro colony-forming activity, and in vivo behavior. We attempted prospective isolation of HSC subsets with distinct properties among CD34(-/low) c-Kit+Sca-1+Lin- (CD34-KSL) cells. CD34-KSL cells were divided, based on CD150 expression, into three fractions: CD150high, CD150med, and CD150neg cells. Compared with the other two fractions, CD150high cells were significantly enriched in HSCs, with great self-renewal potential. In vitro colony assays revealed that decreased expression of CD150 was associated with reduced erythroblast/megakaryocyte differentiation potential. All three fractions were regenerated only from CD150high cells in recipient mice. Using single-cell transplantation studies, we found that a fraction of CD150high cells displayed latent and barely detectable myeloid engraftment in primary-recipient mice but progressive and multilineage reconstitution in secondary-recipient mice. These findings highlight the complexity and hierarchy of reconstitution capability, even among HSCs in the most primitive compartment.
Figures


References
-
- Abkowitz J.L., Golinelli D., Harrison D.E., Guttorp P. 2000. In vivo kinetics of murine hemopoietic stem cells. Blood. 96:3399–3405 - PubMed
-
- Adolfsson J., Borge O.J., Bryder D., Theilgaard-Mönch K., Astrand-Grundström I., Sitnicka E., Sasaki Y., Jacobsen S.E. 2001. Upregulation of Flt3 expression within the bone marrow Lin(−)Sca1(+)c-kit(+) stem cell compartment is accompanied by loss of self-renewal capacity. Immunity. 15:659–669 10.1016/S1074-7613(01)00220-5 - DOI - PubMed
-
- Adolfsson J., Månsson R., Buza-Vidas N., Hultquist A., Liuba K., Jensen C.T., Bryder D., Yang L., Borge O.J., Thoren L.A., et al. 2005. Identification of Flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential: a revised road map for adult blood lineage commitment. Cell. 121:295–306 10.1016/j.cell.2005.02.013 - DOI - PubMed
-
- Arinobu Y., Mizuno S., Chong Y., Shigematsu H., Iino T., Iwasaki H., Graf T., Mayfield R., Chan S., Kastner P., Akashi K. 2007. Reciprocal activation of GATA-1 and PU.1 marks initial specification of hematopoietic stem cells into myeloerythroid and myelolymphoid lineages. Cell Stem Cell. 1:416–427 10.1016/j.stem.2007.07.004 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
