

Proteolysis of human thrombin generates novel host defense peptides
- PMID: 20421939
- PMCID: PMC2858699
- DOI: 10.1371/journal.ppat.1000857
Proteolysis of human thrombin generates novel host defense peptides
Abstract
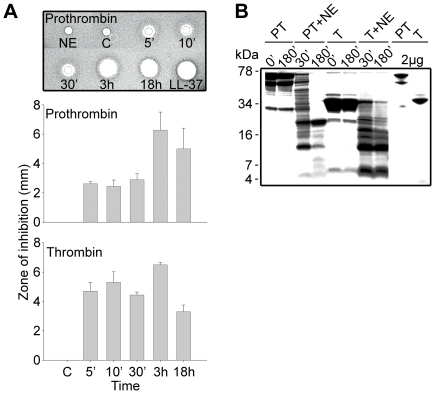
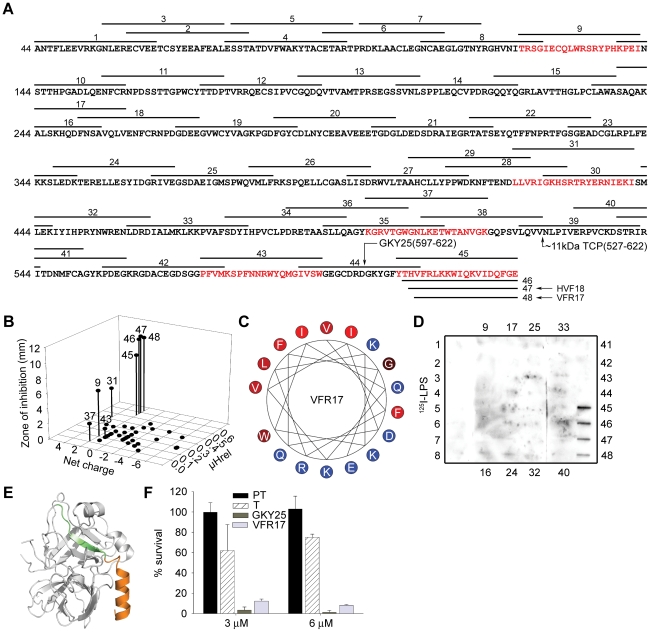
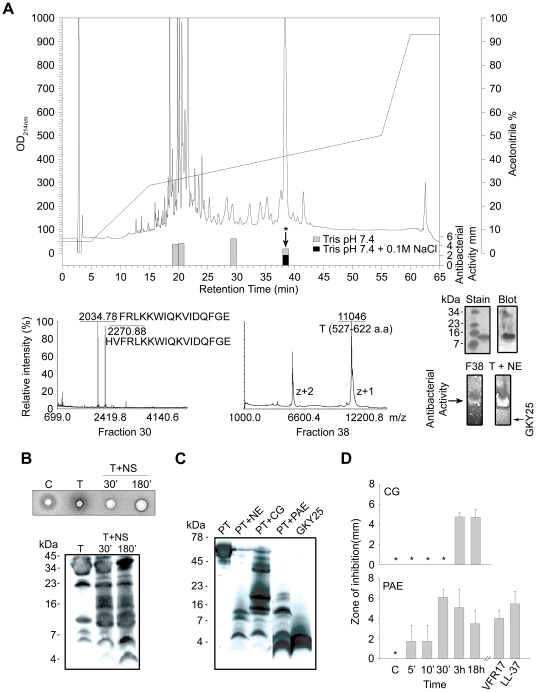
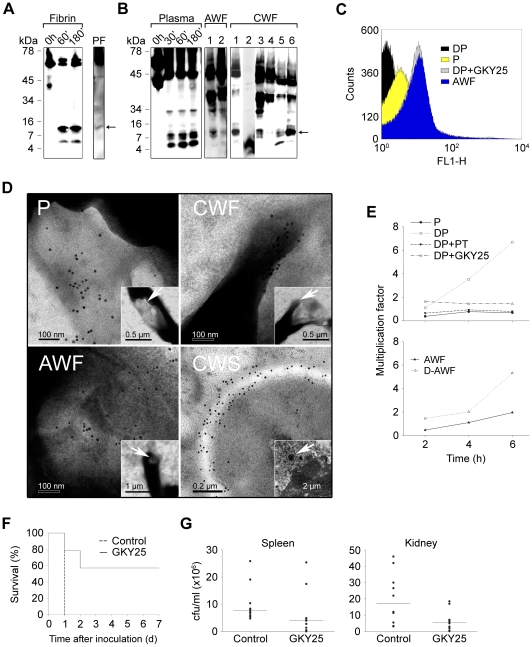
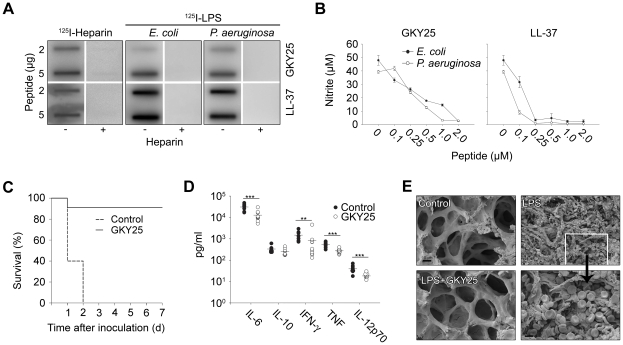
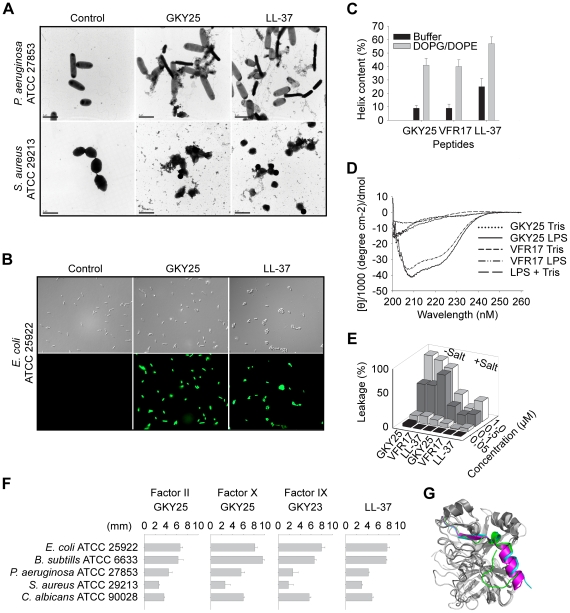
The coagulation system is characterized by the sequential and highly localized activation of a series of serine proteases, culminating in the conversion of fibrinogen into fibrin, and formation of a fibrin clot. Here we show that C-terminal peptides of thrombin, a key enzyme in the coagulation cascade, constitute a novel class of host defense peptides, released upon proteolysis of thrombin in vitro, and detected in human wounds in vivo. Under physiological conditions, these peptides exert antimicrobial effects against Gram-positive and Gram-negative bacteria, mediated by membrane lysis, as well as immunomodulatory functions, by inhibiting macrophage responses to bacterial lipopolysaccharide. In mice, they are protective against P. aeruginosa sepsis, as well as lipopolysaccharide-induced shock. Moreover, the thrombin-derived peptides exhibit helical structures upon binding to lipopolysaccharide and can also permeabilize liposomes, features typical of "classical" helical antimicrobial peptides. These findings provide a novel link between the coagulation system and host-defense peptides, two fundamental biological systems activated in response to injury and microbial invasion.
Conflict of interest statement
Drs. Schmidtchen and Malmsten have shares in DermaGen AB and XImmune AB, companies involved in the therapeutic development of peptides for topical and systemic use, respectively.
Figures
Similar articles
-
The C-terminal sequence of several human serine proteases encodes host defense functions.J Innate Immun. 2011;3(5):471-82. doi: 10.1159/000327016. Epub 2011 May 11. J Innate Immun. 2011. PMID: 21576923
-
Host defense peptides of thrombin modulate inflammation and coagulation in endotoxin-mediated shock and Pseudomonas aeruginosa sepsis.PLoS One. 2012;7(12):e51313. doi: 10.1371/journal.pone.0051313. Epub 2012 Dec 13. PLoS One. 2012. PMID: 23272096 Free PMC article.
-
Aggregation of thrombin-derived C-terminal fragments as a previously undisclosed host defense mechanism.Proc Natl Acad Sci U S A. 2017 May 23;114(21):E4213-E4222. doi: 10.1073/pnas.1619609114. Epub 2017 May 4. Proc Natl Acad Sci U S A. 2017. PMID: 28473418 Free PMC article.
-
Molecular diversity in gene-encoded, cationic antimicrobial polypeptides.Curr Pharm Des. 2002;8(9):743-61. doi: 10.2174/1381612023395475. Curr Pharm Des. 2002. PMID: 11945169 Review.
-
Staphylococcal resistance to antimicrobial peptides of mammalian and bacterial origin.Peptides. 2001 Oct;22(10):1651-9. doi: 10.1016/s0196-9781(01)00500-9. Peptides. 2001. PMID: 11587793 Review.
Cited by
-
Plasma Protein Layer Concealment Protects Streptococcus pyogenes From Innate Immune Attack.Front Cell Infect Microbiol. 2021 May 20;11:633394. doi: 10.3389/fcimb.2021.633394. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34094995 Free PMC article.
-
Selective protein aggregation confines and inhibits endotoxins in wounds: Linking host defense to amyloid formation.iScience. 2023 Sep 19;26(10):107951. doi: 10.1016/j.isci.2023.107951. eCollection 2023 Oct 20. iScience. 2023. PMID: 37817942 Free PMC article.
-
Neutrophil serine proteases in antibacterial defense.Curr Opin Microbiol. 2015 Feb;23:42-8. doi: 10.1016/j.mib.2014.11.002. Epub 2014 Nov 18. Curr Opin Microbiol. 2015. PMID: 25461571 Free PMC article. Review.
-
Immunohaemostasis: a new view on haemostasis during sepsis.Ann Intensive Care. 2017 Dec 2;7(1):117. doi: 10.1186/s13613-017-0339-5. Ann Intensive Care. 2017. PMID: 29197958 Free PMC article. Review.
-
Proteomic Screening for Prediction and Design of Antimicrobial Peptides with AmpGram.Int J Mol Sci. 2020 Jun 17;21(12):4310. doi: 10.3390/ijms21124310. Int J Mol Sci. 2020. PMID: 32560350 Free PMC article.
References
-
- Lehrer RI, Ganz T. Cathelicidins: a family of endogenous antimicrobial peptides. Curr Opin Hematol. 2002;9:18–22. - PubMed
-
- Harder J, Glaser R, Schröder JM. Review: Human antimicrobial proteins effectors of innate immunity. J Endotoxin Res. 2007;13:317–338. - PubMed
-
- Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415:389–395. - PubMed
-
- Tossi A, Sandri L, Giangaspero A. Amphipathic, alpha-helical antimicrobial peptides. Biopolymers. 2000;55:4–30. - PubMed
-
- Yount NY, Bayer AS, Xiong YQ, Yeaman MR. Advances in antimicrobial peptide immunobiology. Biopolymers. 2006;84:435–458. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
