

The type I NADH dehydrogenase of Mycobacterium tuberculosis counters phagosomal NOX2 activity to inhibit TNF-alpha-mediated host cell apoptosis
- PMID: 20421951
- PMCID: PMC2858756
- DOI: 10.1371/journal.ppat.1000864
The type I NADH dehydrogenase of Mycobacterium tuberculosis counters phagosomal NOX2 activity to inhibit TNF-alpha-mediated host cell apoptosis
Abstract
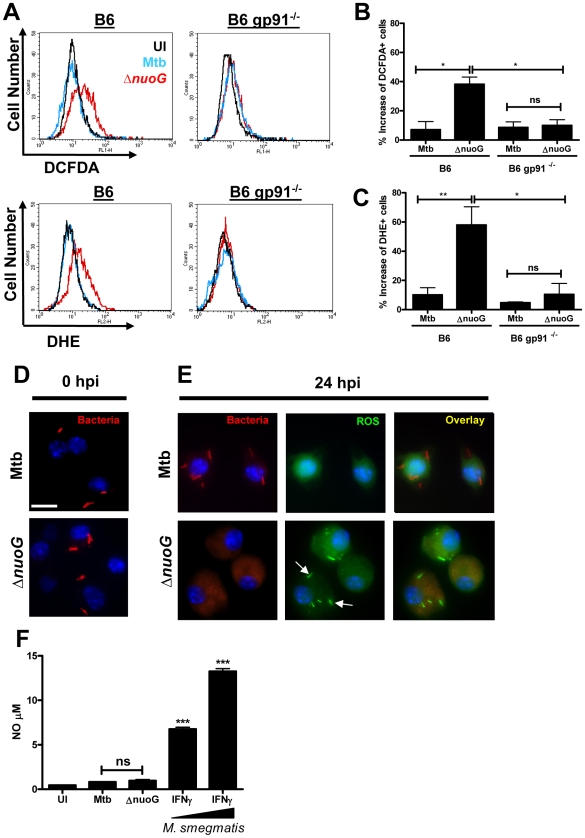
The capacity of infected cells to undergo apoptosis upon insult with a pathogen is an ancient innate immune defense mechanism. Consequently, the ability of persisting, intracellular pathogens such as the human pathogen Mycobacterium tuberculosis (Mtb) to inhibit infection-induced apoptosis of macrophages is important for virulence. The nuoG gene of Mtb, which encodes the NuoG subunit of the type I NADH dehydrogenase, NDH-1, is important in Mtb-mediated inhibition of host macrophage apoptosis, but the molecular mechanism of this host pathogen interaction remains elusive. Here we show that the apoptogenic phenotype of MtbDeltanuoG was significantly reduced in human macrophages treated with caspase-3 and -8 inhibitors, TNF-alpha-neutralizing antibodies, and also after infection of murine TNF(-/-) macrophages. Interestingly, incubation of macrophages with inhibitors of reactive oxygen species (ROS) reduced not only the apoptosis induced by the nuoG mutant, but also its capacity to increase macrophage TNF-alpha secretion. The MtbDeltanuoG phagosomes showed increased ROS levels compared to Mtb phagosomes in primary murine and human alveolar macrophages. The increase in MtbDeltanuoG induced ROS and apoptosis was abolished in NOX-2 deficient (gp91(-/-)) macrophages. These results suggest that Mtb, via a NuoG-dependent mechanism, can neutralize NOX2-derived ROS in order to inhibit TNF-alpha-mediated host cell apoptosis. Consistently, an Mtb mutant deficient in secreted catalase induced increases in phagosomal ROS and host cell apoptosis, both of which were dependent upon macrophage NOX-2 activity. In conclusion, these results serendipitously reveal a novel connection between NOX2 activity, phagosomal ROS, and TNF-alpha signaling during infection-induced apoptosis in macrophages. Furthermore, our study reveals a novel function of NOX2 activity in innate immunity beyond the initial respiratory burst, which is the sensing of persistent intracellular pathogens and subsequent induction of host cell apoptosis as a second line of defense.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






Similar articles
-
NADPH oxidase 2 interaction with TLR2 is required for efficient innate immune responses to mycobacteria via cathelicidin expression.J Immunol. 2009 Mar 15;182(6):3696-705. doi: 10.4049/jimmunol.0802217. J Immunol. 2009. PMID: 19265148
-
Mycobacterium tuberculosis nucleoside diphosphate kinase inactivates small GTPases leading to evasion of innate immunity.PLoS Pathog. 2013;9(7):e1003499. doi: 10.1371/journal.ppat.1003499. Epub 2013 Jul 18. PLoS Pathog. 2013. PMID: 23874203 Free PMC article.
-
Ligation of FcγR Alters Phagosomal Processing of Protein via Augmentation of NADPH Oxidase Activity.Traffic. 2016 Jul;17(7):786-802. doi: 10.1111/tra.12396. Epub 2016 May 13. Traffic. 2016. PMID: 27020146
-
Host Cell Death and Modulation of Immune Response against Mycobacterium tuberculosis Infection.Int J Mol Sci. 2024 Jun 6;25(11):6255. doi: 10.3390/ijms25116255. Int J Mol Sci. 2024. PMID: 38892443 Free PMC article. Review.
-
Host Cell Targets of Released Lipid and Secreted Protein Effectors of Mycobacterium tuberculosis.Front Cell Infect Microbiol. 2020 Oct 23;10:595029. doi: 10.3389/fcimb.2020.595029. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33194845 Free PMC article. Review.
Cited by
-
A rho GDP dissociation inhibitor produced by apoptotic T-cells inhibits growth of Mycobacterium tuberculosis.PLoS Pathog. 2015 Feb 6;11(2):e1004617. doi: 10.1371/journal.ppat.1004617. eCollection 2015 Feb. PLoS Pathog. 2015. PMID: 25659138 Free PMC article.
-
A secreted form of chorismate mutase (Rv1885c) in Mycobacterium bovis BCG contributes to pathogenesis by inhibiting mitochondria-mediated apoptotic cell death of macrophages.J Biomed Sci. 2023 Dec 18;30(1):95. doi: 10.1186/s12929-023-00988-2. J Biomed Sci. 2023. PMID: 38110948 Free PMC article.
-
Suboptimal activation of antigen-specific CD4+ effector cells enables persistence of M. tuberculosis in vivo.PLoS Pathog. 2011 May;7(5):e1002063. doi: 10.1371/journal.ppat.1002063. Epub 2011 May 26. PLoS Pathog. 2011. PMID: 21637811 Free PMC article.
-
Intelligent Mechanisms of Macrophage Apoptosis Subversion by Mycobacterium.Pathogens. 2020 Mar 16;9(3):218. doi: 10.3390/pathogens9030218. Pathogens. 2020. PMID: 32188164 Free PMC article. Review.
-
Mycobacterium tuberculosis Rv3402c enhances mycobacterial survival within macrophages and modulates the host pro-inflammatory cytokines production via NF-kappa B/ERK/p38 signaling.PLoS One. 2014 Apr 10;9(4):e94418. doi: 10.1371/journal.pone.0094418. eCollection 2014. PLoS One. 2014. PMID: 24722253 Free PMC article.
References
-
- Bedard K, Krause KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev. 2007;87:245–313. - PubMed
-
- Fang FC. Antimicrobial reactive oxygen and nitrogen species: concepts and controversies. Nat Rev Microbiol. 2004;2:820–832. - PubMed
-
- Savina A, Amigorena S. Phagocytosis and antigen presentation in dendritic cells. Immunol Rev. 2007;219:143–156. - PubMed
-
- Freeman AF, Holland SM. Persistent bacterial infections and primary immune disorders. Curr Opin Microbiol. 2007;10:70–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
