

The potential of human allogeneic juvenile chondrocytes for restoration of articular cartilage
- PMID: 20423988
- PMCID: PMC3774103
- DOI: 10.1177/0363546510361950
The potential of human allogeneic juvenile chondrocytes for restoration of articular cartilage
Abstract
Background: Donor-site morbidity, limited numbers of cells, loss of phenotype during ex vivo expansion, and age-related decline in chondrogenic activity present critical obstacles to the use of autologous chondrocyte implantation for cartilage repair. Chondrocytes from juvenile cadaveric donors may represent an alternative to autologous cells. Hypothesis/
Purpose: The authors hypothesized that juvenile chondrocyte would show stronger and more stable chondrogenic activity than adult cells in vitro and that juvenile cells pose little risk of immunologic incompatibility in adult hosts.
Study design: Controlled laboratory study.
Methods: Cartilage samples were from juvenile (<13 years old) and adult (>13 years old) donors. The chondrogenic activity of freshly isolated human articular chondrocytes and of expanded cells after monolayer culture was measured by proteoglycan assay, gene expression analysis, and histology. Lymphocyte proliferation assays were used to assess immunogenic activity.
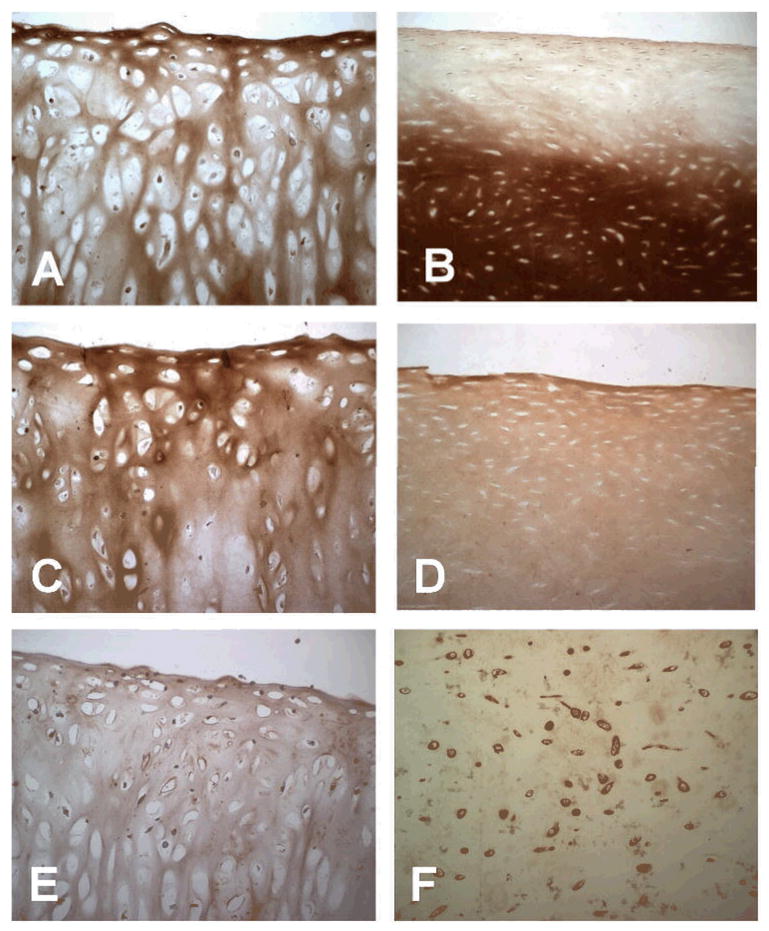
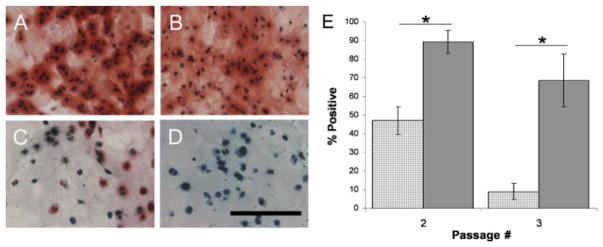
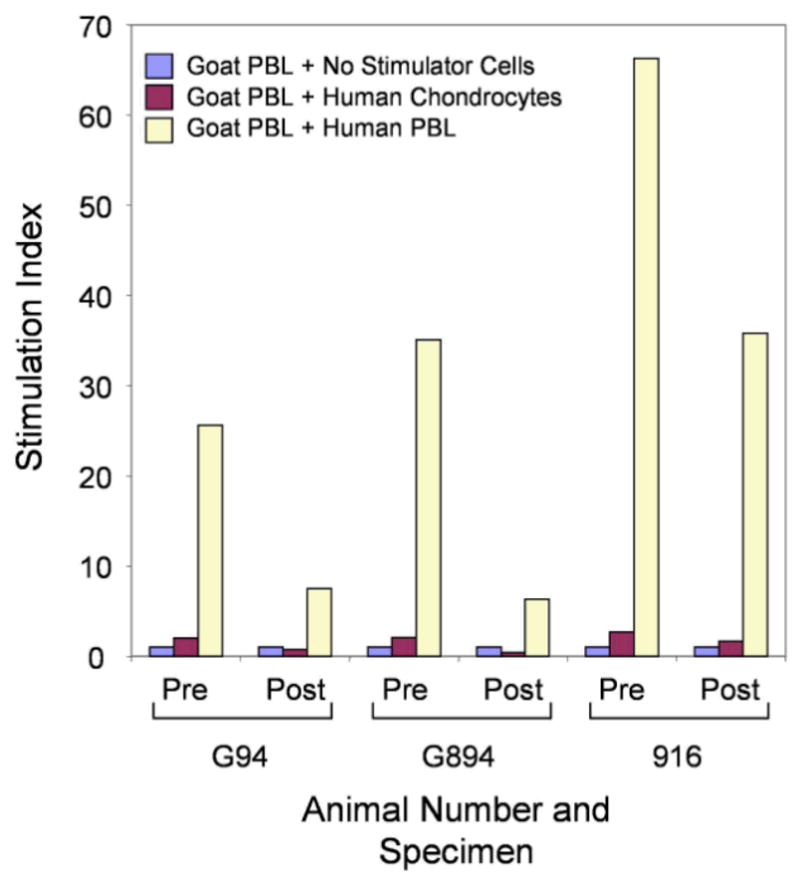
Results: Proteoglycan content in neocartilage produced by juvenile chondrocytes was 100-fold higher than in neocartilage produced by adult cells. Collagen type II and type IX mRNA in fresh juvenile chondrocytes were 100- and 700-fold higher, respectively, than in adult chondrocytes. The distributions of collagens II and IX were similar in native juvenile cartilage and in neocartilage made by juvenile cells. Juvenile cells grew significantly faster in monolayer cultures than adult cells (P = .002) and proteoglycan levels produced in agarose culture was significantly higher in juvenile cells than in adult cells after multiple passages (P < .001). Juvenile chondrocytes did not stimulate lymphocyte proliferation.
Conclusion: These results document a dramatic age-related decline in human chondrocyte chondrogenic potential and show that allogeneic juvenile chondrocytes do not stimulate an immunologic response in vivo.
Clinical relevance: Juvenile human chondrocytes have greater potential to restore articular cartilage than adult cells, and may be transplanted without the fear of rejection, suggesting a new allogeneic approach to restoring articular cartilage in older individuals.
Figures




References
-
- Adkisson HD, Gillis MP, Davis EC, Maloney W, Hruska KA. In vitro generation of scaffold independent neocartilage. Clin Orthop Relat Res. 2001 Oct;(391 Suppl):S280–294. - PubMed
-
- Adkisson HD, Milliman C, Zhang X, Mauch K, Maziarz RT, Streeter PR. Immune evasion by neocartilage derived articular chondrocytes. Stem Cell Res. 2009 Accepted. - PubMed
-
- Aigner T, Gebhard PM, Schmid E, Bau B, Harley V, Poschl E. SOX9 expression does not correlate with type II collagen expression in adult articular chondrocytes. Matrix Biol. 2003 Jun;22(4):363–372. - PubMed
-
- Aston JE, Bentley G. Repair of articular surfaces by allografts of articular and growth-plate cartilage. J Bone Joint Surg Br. 1986 Jan;68(1):29–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
