

Sulfite reductase defines a newly discovered bottleneck for assimilatory sulfate reduction and is essential for growth and development in Arabidopsis thaliana
- PMID: 20424176
- PMCID: PMC2879758
- DOI: 10.1105/tpc.110.074088
Sulfite reductase defines a newly discovered bottleneck for assimilatory sulfate reduction and is essential for growth and development in Arabidopsis thaliana
Abstract
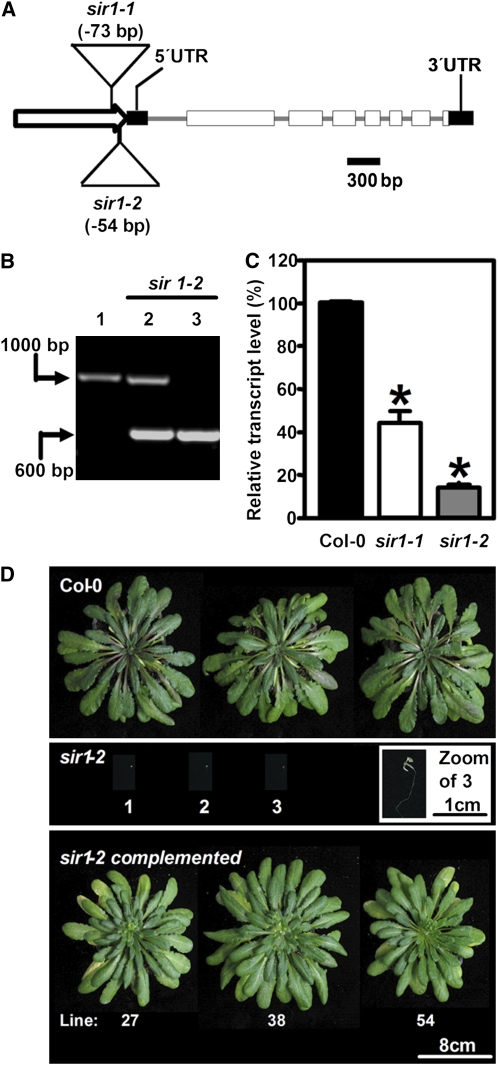
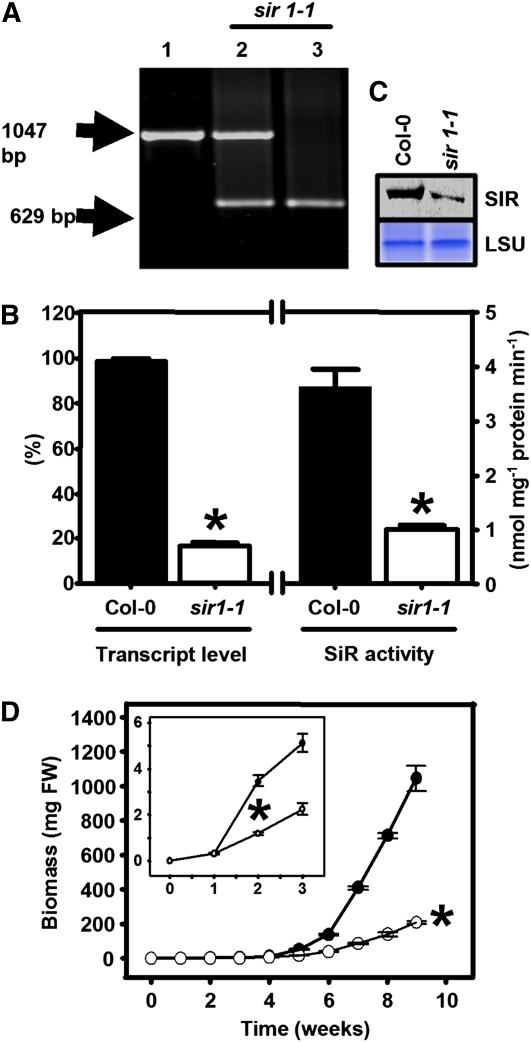
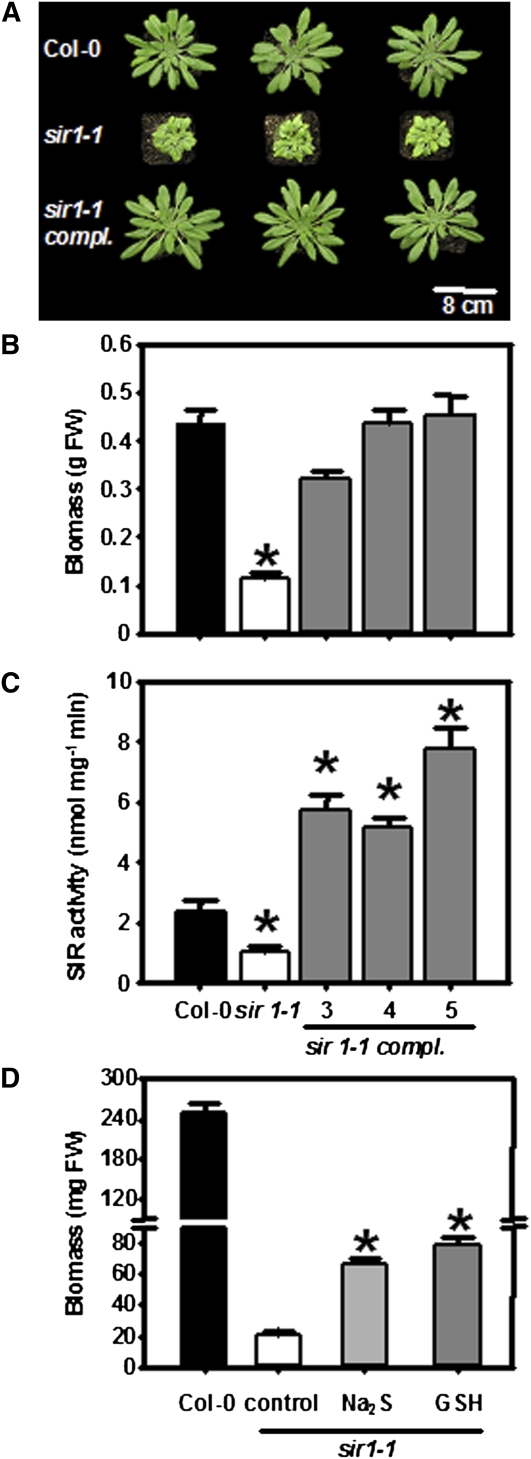
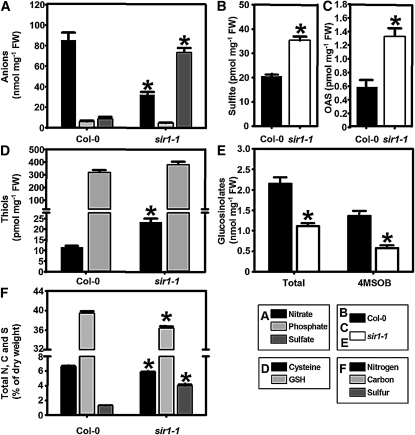
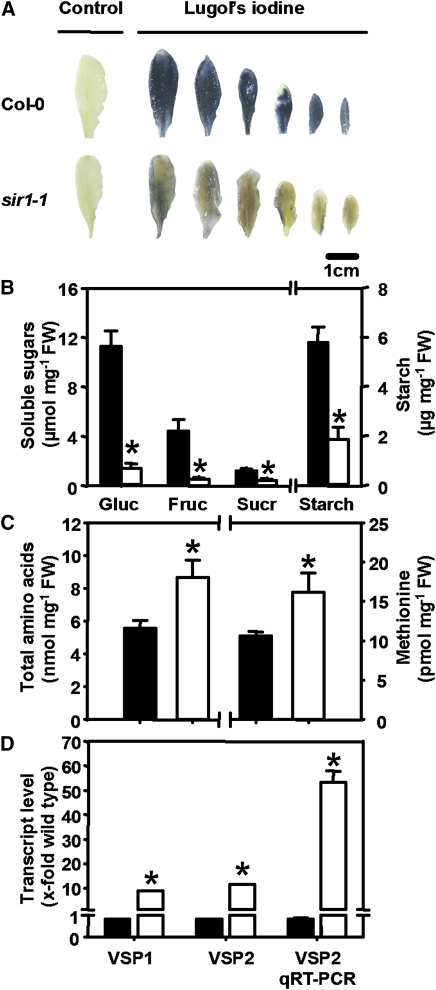
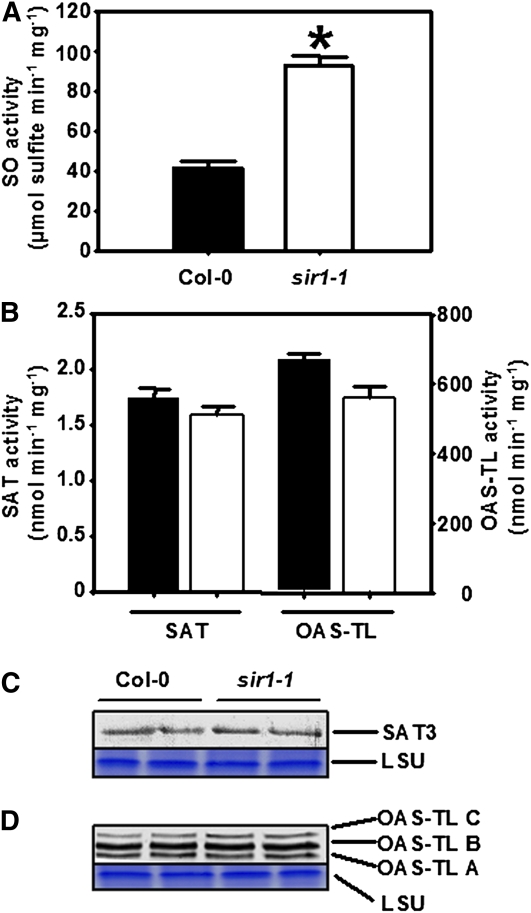
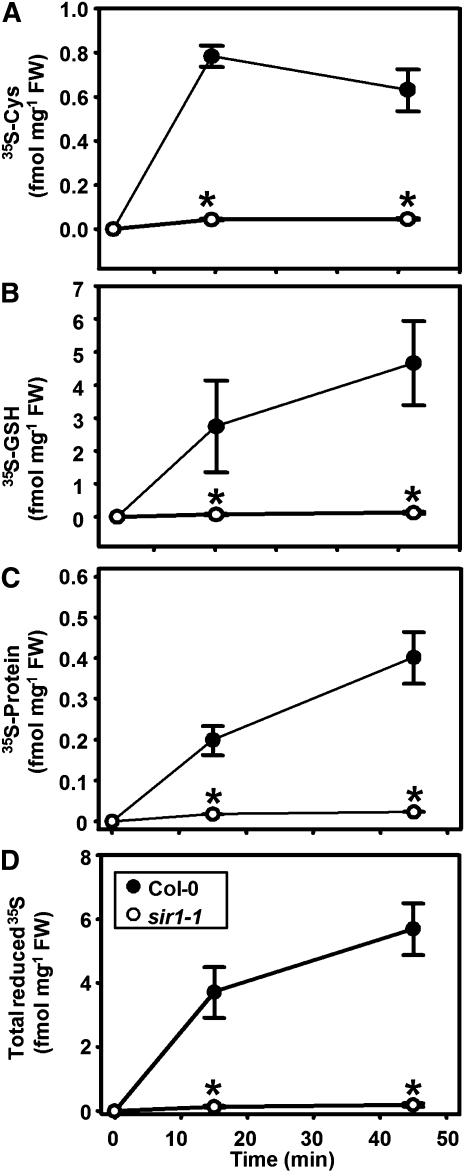
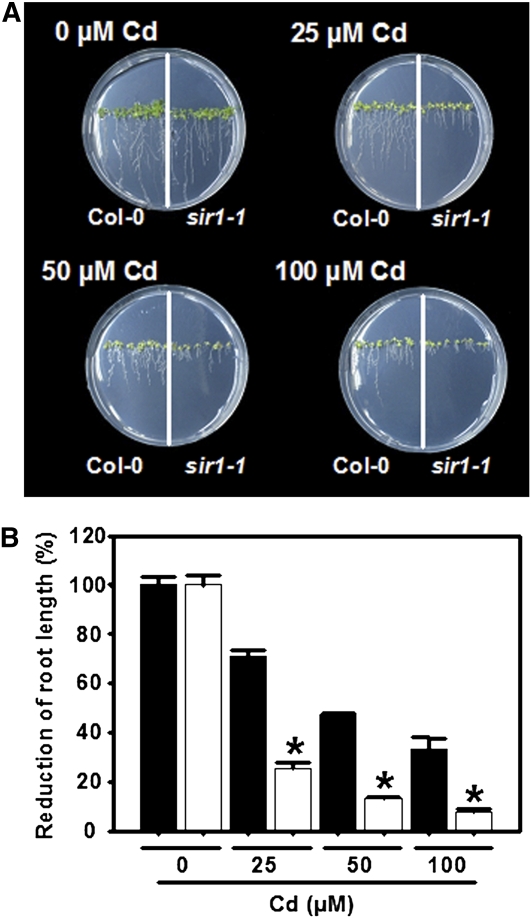
The role of sulfite reductase (SiR) in assimilatory reduction of inorganic sulfate to sulfide has long been regarded as insignificant for control of flux in this pathway. Two independent Arabidopsis thaliana T-DNA insertion lines (sir1-1 and sir1-2), each with an insertion in the promoter region of SiR, were isolated. sir1-2 seedlings had 14% SiR transcript levels compared with the wild type and were early seedling lethal. sir1-1 seedlings had 44% SiR transcript levels and were viable but strongly retarded in growth. In mature leaves of sir1-1 plants, the levels of SiR transcript, protein, and enzymatic activity ranged between 17 and 28% compared with the wild type. The 28-fold decrease of incorporation of (35)S label into Cys, glutathione, and protein in sir1-1 showed that the decreased activity of SiR generated a severe bottleneck in the assimilatory sulfate reduction pathway. Root sulfate uptake was strongly enhanced, and steady state levels of most of the sulfur-related metabolites, as well as the expression of many primary metabolism genes, were changed in leaves of sir1-1. Hexose and starch contents were decreased, while free amino acids increased. Inorganic carbon, nitrogen, and sulfur composition was also severely altered, demonstrating strong perturbations in metabolism that differed markedly from known sulfate deficiency responses. The results support that SiR is the only gene with this function in the Arabidopsis genome, that optimal activity of SiR is essential for normal growth, and that its downregulation causes severe adaptive reactions of primary and secondary metabolism.
Figures

References
-
- Benjamini Y., Hochberg Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc., B 57: 289–300
-
- Benning C. (2007). Questions remaining in sulfolipid biosynthesis: A historical perspective. Photosynth. Res. 92: 199–203 - PubMed
-
- Bork C., Schwenn J.D., Hell R. (1998). Isolation and characterization of a gene for assimilatory sulfite reductase from Arabidopsis thaliana. Gene 212: 147–153 - PubMed
-
- Bradford M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72: 248–254 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
