

Heat generates oxidized linoleic acid metabolites that activate TRPV1 and produce pain in rodents
- PMID: 20424317
- PMCID: PMC2860941
- DOI: 10.1172/JCI41678
Heat generates oxidized linoleic acid metabolites that activate TRPV1 and produce pain in rodents
Abstract
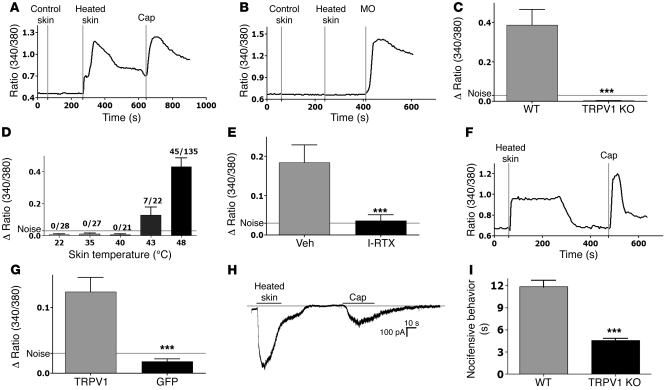
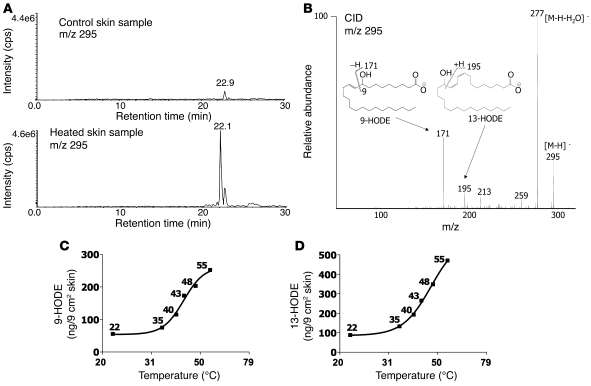
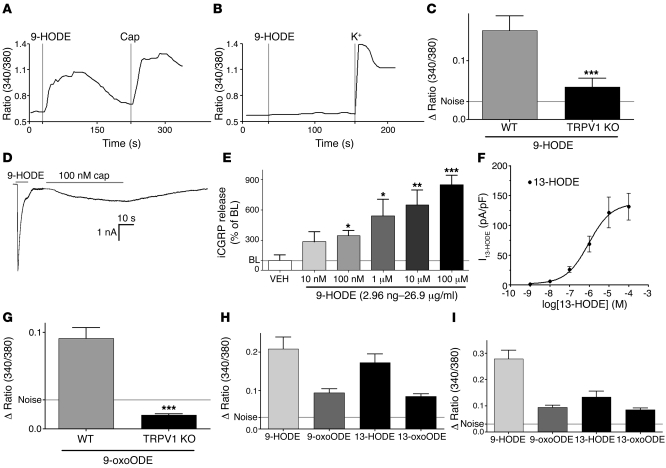
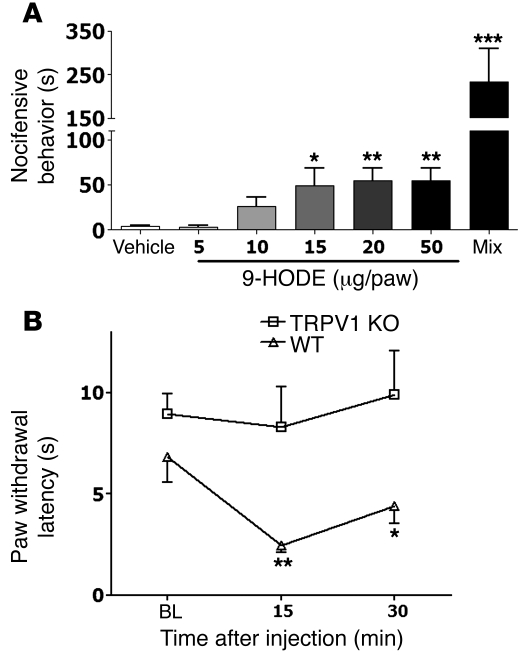
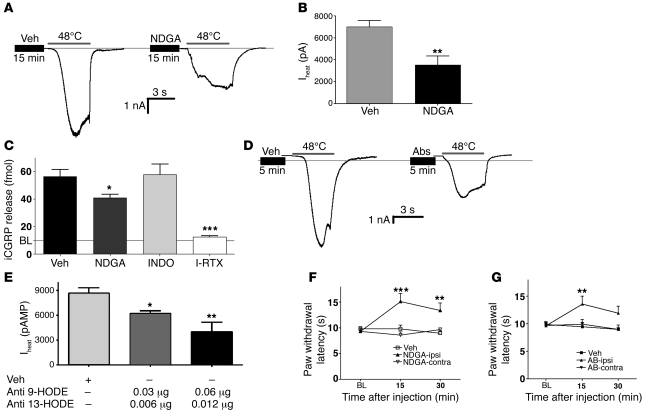
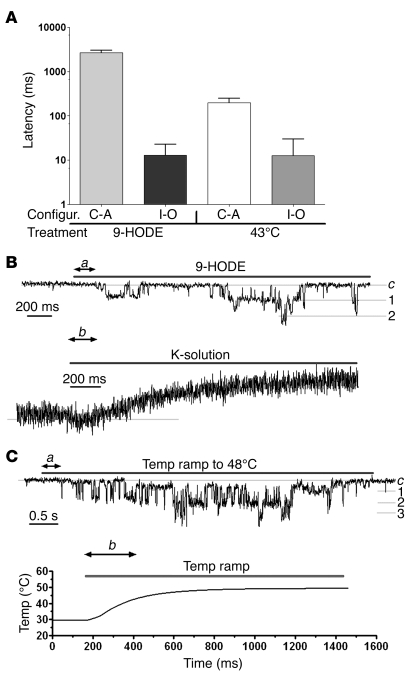
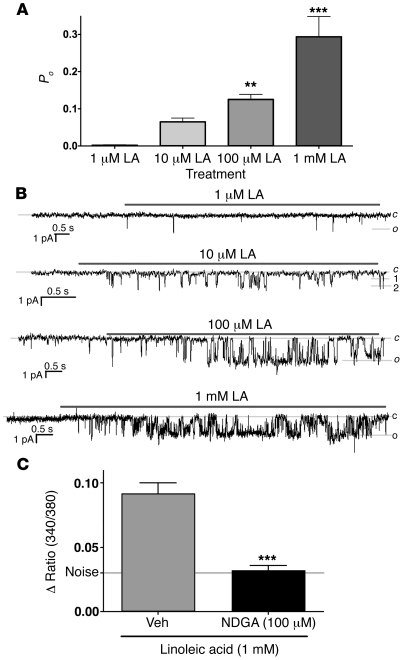
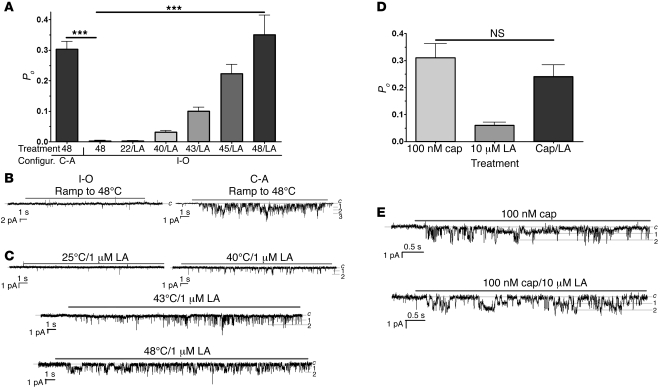
The transient receptor potential vanilloid 1 (TRPV1) channel is the principal detector of noxious heat in the peripheral nervous system. TRPV1 is expressed in many nociceptors and is involved in heat-induced hyperalgesia and thermoregulation. The precise mechanism or mechanisms mediating the thermal sensitivity of TRPV1 are unknown. Here, we have shown that the oxidized linoleic acid metabolites 9- and 13-hydroxyoctadecadienoic acid (9- and 13-HODE) are formed in mouse and rat skin biopsies by exposure to noxious heat. 9- and 13-HODE and their metabolites, 9- and 13-oxoODE, activated TRPV1 and therefore constitute a family of endogenous TRPV1 agonists. Moreover, blocking these substances substantially decreased the heat sensitivity of TRPV1 in rats and mice and reduced nociception. Collectively, our results indicate that HODEs contribute to the heat sensitivity of TRPV1 in rodents. Because oxidized linoleic acid metabolites are released during cell injury, these findings suggest a mechanism for integrating the hyperalgesic and proinflammatory roles of TRPV1 and linoleic acid metabolites and may provide the foundation for investigating new classes of analgesic drugs.
Figures
Comment in
-
Some new insights into the molecular mechanisms of pain perception.J Clin Invest. 2010 May;120(5):1380-3. doi: 10.1172/JCI42143. Epub 2010 Apr 26. J Clin Invest. 2010. PMID: 20424321 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
