

Peroxynitrite toxicity in Escherichia coli K12 elicits expression of oxidative stress responses and protein nitration and nitrosylation
- PMID: 20427277
- PMCID: PMC2898335
- DOI: 10.1074/jbc.M109.085506
Peroxynitrite toxicity in Escherichia coli K12 elicits expression of oxidative stress responses and protein nitration and nitrosylation
Abstract
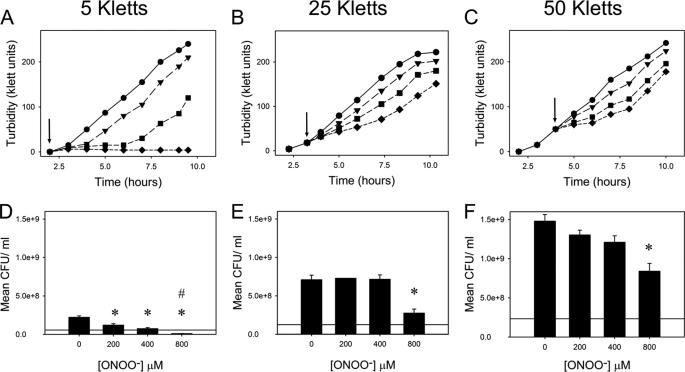
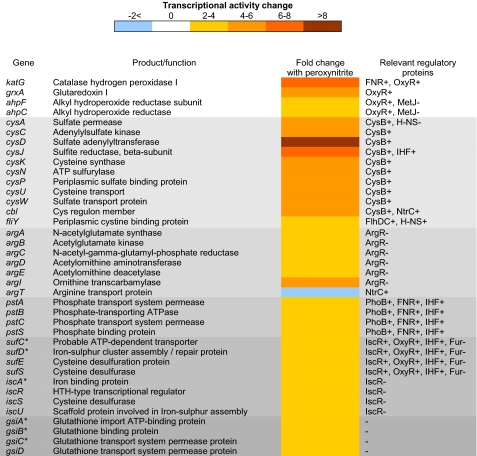
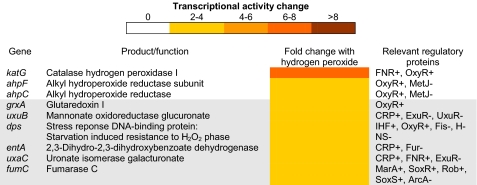
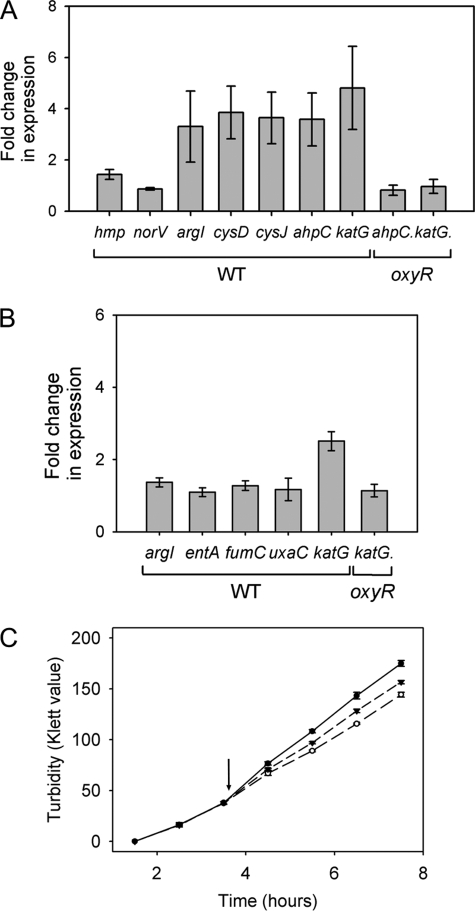
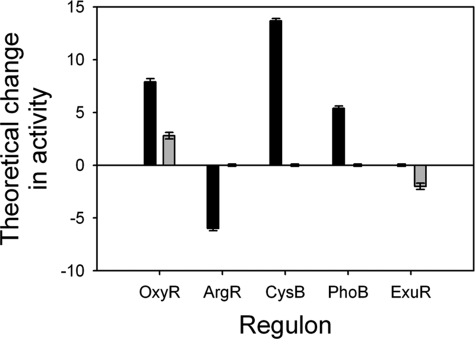
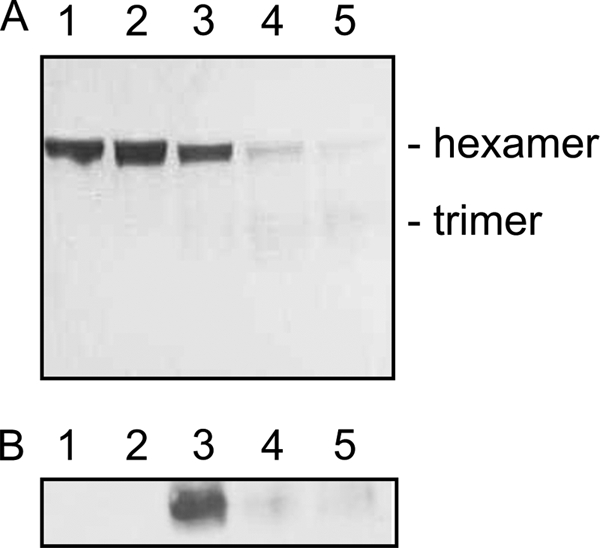
Peroxynitrite is formed in macrophages by the diffusion-limited reaction of superoxide and nitric oxide. This highly reactive species is thought to contribute to bacterial killing by interaction with diverse targets and nitration of protein tyrosines. This work presents for the first time a comprehensive analysis of transcriptional responses to peroxynitrite under tightly controlled chemostat growth conditions. Up-regulation of the cysteine biosynthesis pathway and an increase in S-nitrosothiol levels suggest S-nitrosylation to be a consequence of peroxynitrite exposure. Genes involved in the assembly/repair of iron-sulfur clusters also show enhanced transcription. Unexpectedly, arginine biosynthesis gene transcription levels were also elevated after treatment with peroxynitrite. Analysis of the negative regulator for these genes, ArgR, showed that post-translational nitration of tyrosine residues within this protein is responsible for its degradation in vitro. Further up-regulation was seen in oxidative stress response genes, including katG and ahpCF. However, genes known to be up-regulated by nitric oxide and nitrosating agents (e.g. hmp and norVW) were unaffected. Probabilistic modeling of the transcriptomic data identified five altered transcription factors in response to peroxynitrite exposure, including OxyR and ArgR. Hydrogen peroxide can be present as a contaminant in commercially available peroxynitrite preparations. Transcriptomic analysis of cells treated with hydrogen peroxide alone also revealed up-regulation of oxidative stress response genes but not of many other genes that are up-regulated by peroxynitrite. Thus, the cellular responses to peroxynitrite and hydrogen peroxide are distinct.
Figures
References
-
- Bryk R., Griffin P., Nathan C. (2000) Nature 407, 211–215 - PubMed
-
- Master S. S., Springer B., Sander P., Boettger E. C., Deretic V., Timmins G. S. (2002) Microbiology 148, 3139–3144 - PubMed
-
- Pfeiffer S., Gorren A. C., Schmidt K., Werner E. R., Hansert B., Bohle D. S., Mayer B. (1997) J. Biol. Chem. 272, 3465–3470 - PubMed
-
- Ischiropoulos H. (2009) Arch. Biochem. Biophys. 484, 117–121 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
