

The evolution of thrombospondins and their ligand-binding activities
- PMID: 20427418
- PMCID: PMC3107593
- DOI: 10.1093/molbev/msq107
The evolution of thrombospondins and their ligand-binding activities
Abstract
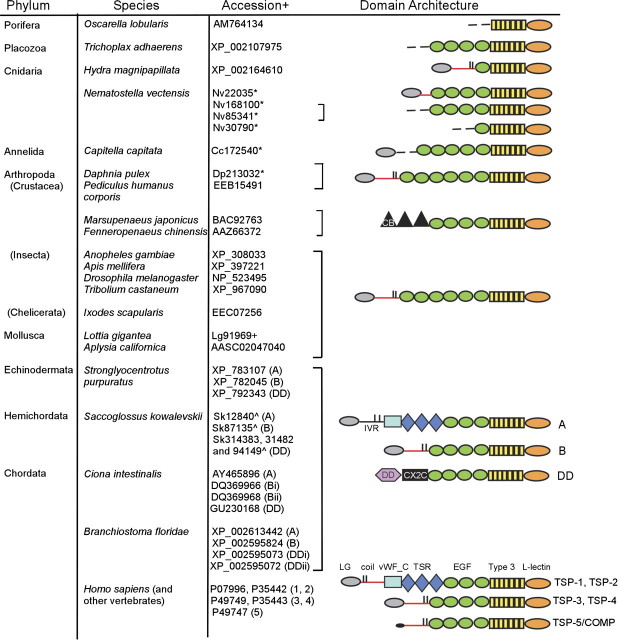
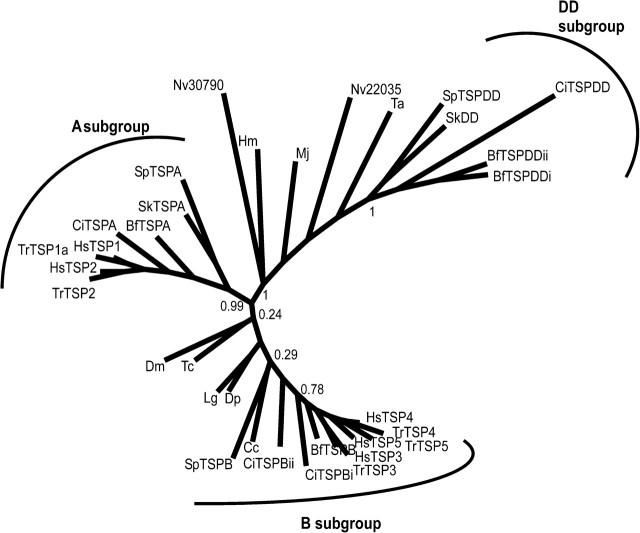
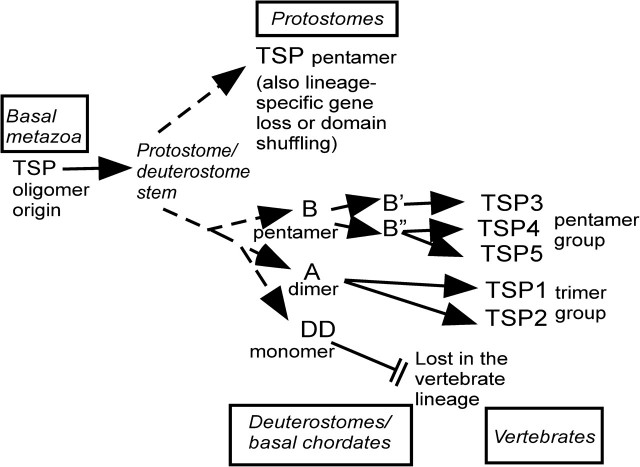
The extracellular matrix (ECM) is a complex, multiprotein network that has essential roles in tissue integrity and intercellular signaling in the metazoa. Thrombospondins (TSPs) are extracellular, calcium-binding glycoproteins that have biologically important roles in mammals in angiogenesis, vascular biology, connective tissues, immune response, and synaptogenesis. The evolution of these complex functional properties is poorly understood. We report here on the evolution of TSPs and their ligand-binding capacities, from comparative genomics of species representing the major phyla of metazoa and experimental analyses of the oligomerization properties of noncanonical TSPs of basal deuterostomes. Monomeric, dimeric, trimeric, and pentameric TSPs have arisen through separate evolutionary events involving gain, loss, or modification of a coiled-coil domain or distinct domains at the amino-terminus. The relative transience of monomeric forms under evolution implicates a biological importance for multivalency of the C-terminal region of TSPs. Most protostomes have a single TSP gene encoding a pentameric TSP. The pentameric form is also present in deuterostomes, and gene duplications at the origin of deuterostomes and gene loss and further gene duplication events in the vertebrate lineage gave rise to distinct forms and novel domain architectures. Parallel analysis of the major ligands of mammalian TSPs revealed that many binding activities are neofunctions representing either coevolutionary innovations in the deuterostome lineage or neofunctions of ancient molecules such as CD36. Contrasting widely conserved capacities include binding to heparan glycosaminoglycans, fibrillar collagen, or RGD-dependent integrins. These findings identify TSPs as fundamental components of the extracellular interaction systems of metazoa and thus impact understanding of the evolution of ECM networks. The widely conserved activities of TSPs in binding to ECM components or PS2 clade integrins will be relevant to use of TSPs in synthetic extracellular matrices or tissue engineering. In contrast, the neofunctions of vertebrate TSPs likely include interactions suitable for therapeutic targeting without general disruption of ECM.
Figures


References
-
- Adams JC. Thrombospondins: multifunctional regulators of cell interactions. Annu Rev Cell Dev Biol. 2001;17:25–51. - PubMed
-
- Adams JC. Functions of the conserved thrombospondin carboxy-terminal cassette in cell-extracellular matrix interactions and signaling. Int J Biochem Cell Biol. 2004;36:1102–1114. - PubMed
-
- Adams JC, Bentley AA, Kvansakul M, Hatherley D, Hohenester E. Extracellular matrix retention of thrombospondin 1 is controlled by its conserved C-terminal region. J Cell Sci. 2008;121:784–795. - PubMed
-
- Adams JC, Monk R, Taylor AL, Ozbek S, Fascetti N, Baumgartner S, Engel J. Characterisation of Drosophila thrombospondin defines an early origin of pentameric thrombospondins. J Mol Biol. 2003;328:479–494. - PubMed
-
- Adams MD, Celniker SE, Holt RA, et al. (195 co-authors) The genome sequence of Drosophila melanogaster. Science. 2000;287:2185–2195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
