

Brain-derived neurotrophic factor regulates satellite cell differentiation and skeltal muscle regeneration
- PMID: 20427568
- PMCID: PMC2893983
- DOI: 10.1091/mbc.e10-02-0154
Brain-derived neurotrophic factor regulates satellite cell differentiation and skeltal muscle regeneration
Abstract
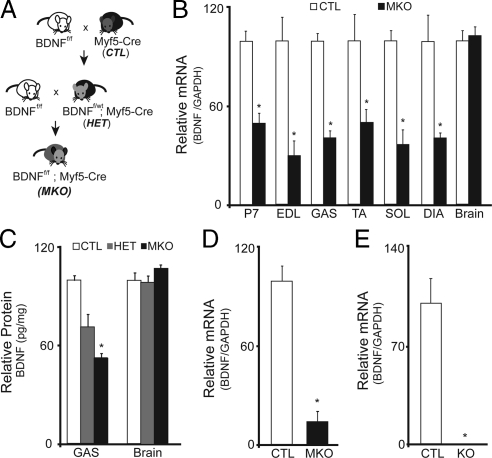
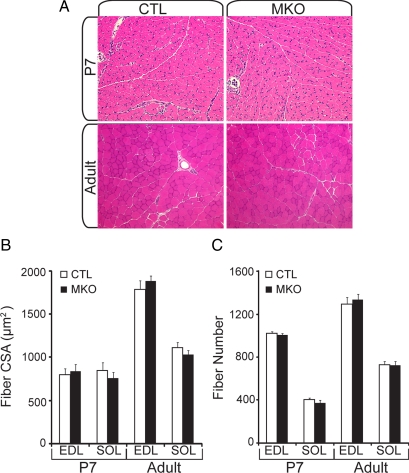
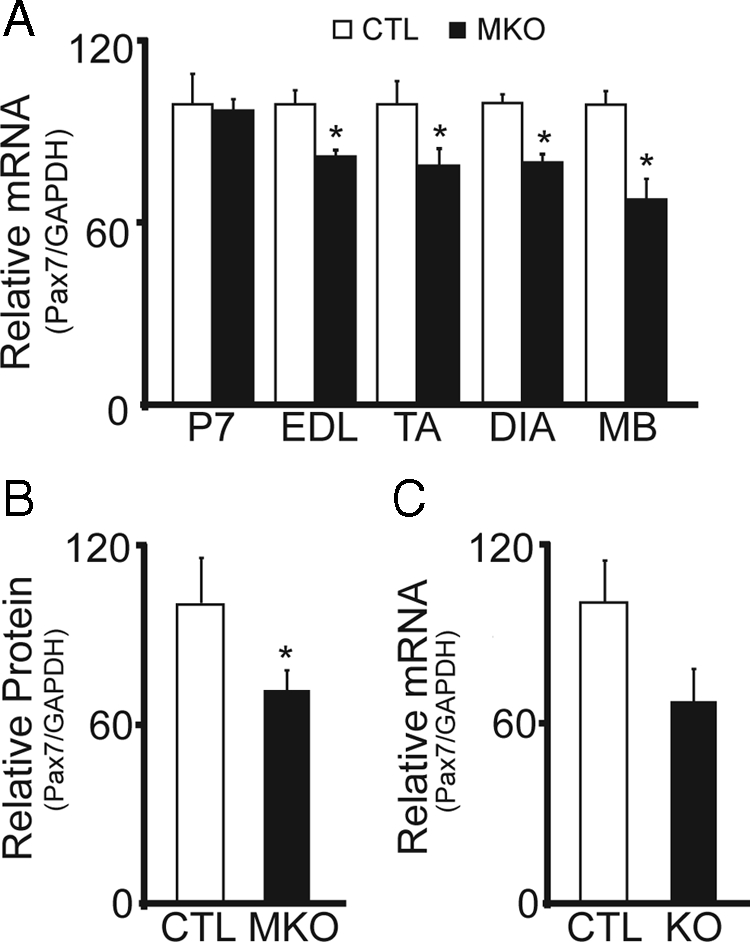
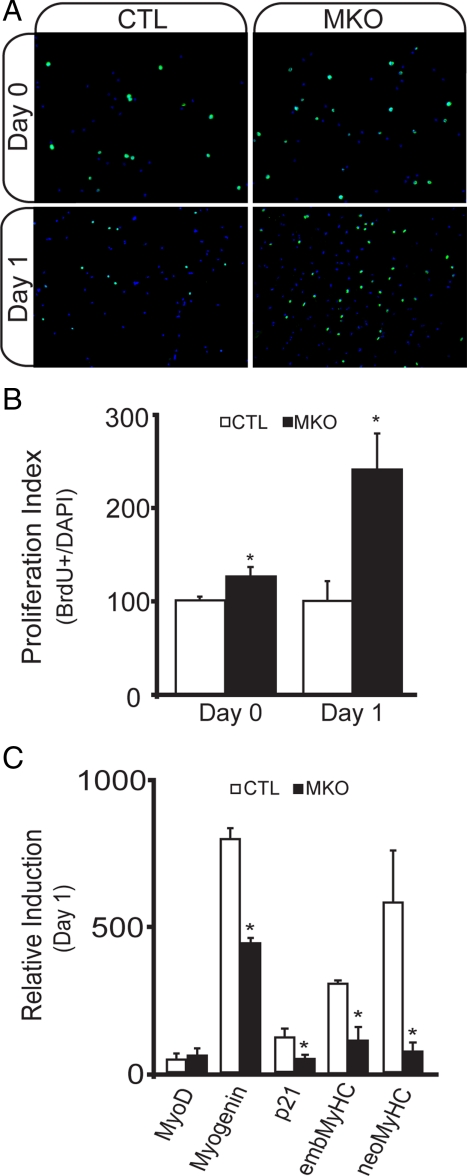
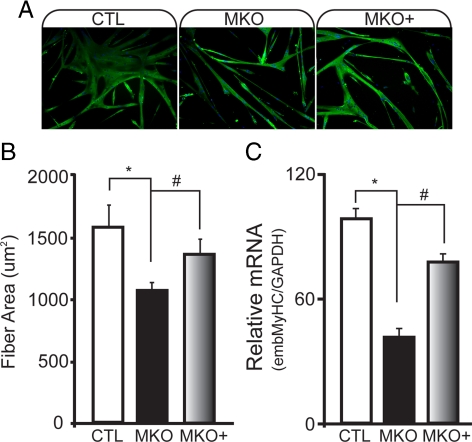
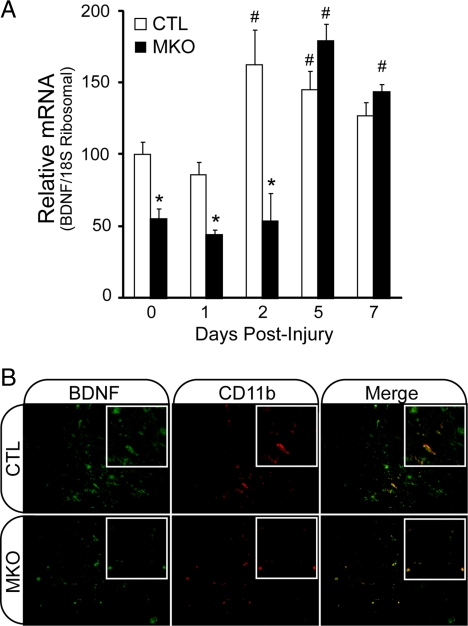
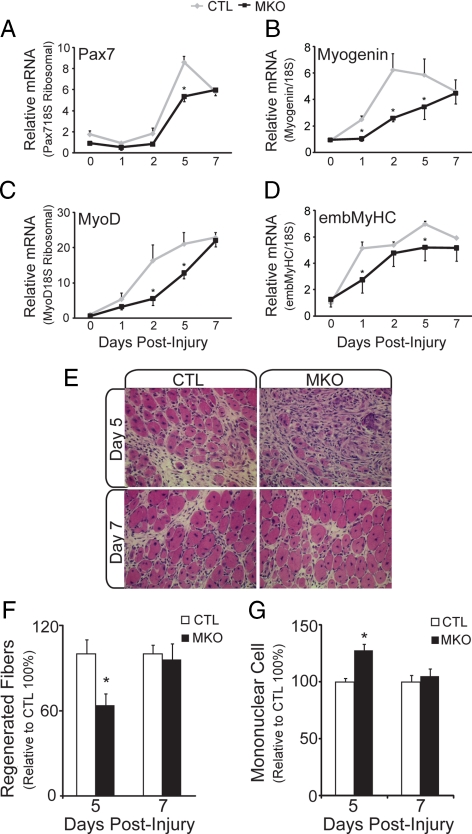
In adult skeletal muscle, brain-derived neurotrophic factor (BDNF) is expressed in myogenic progenitors known as satellite cells. To functionally address the role of BDNF in muscle satellite cells and regeneration in vivo, we generated a mouse in which BDNF is specifically depleted from skeletal muscle cells. For comparative purposes, and to determine the specific role of muscle-derived BDNF, we also examined muscles of the complete BDNF(-/-) mouse. In both models, expression of the satellite cell marker Pax7 was significantly decreased. Furthermore, proliferation and differentiation of primary myoblasts was abnormal, exhibiting delayed induction of several markers of differentiation as well as decreased myotube size. Treatment with exogenous BDNF protein was sufficient to rescue normal gene expression and myotube size. Because satellite cells are responsible for postnatal growth and repair of skeletal muscle, we next examined whether regenerative capacity was compromised. After injury, BDNF-depleted muscle showed delayed expression of several molecular markers of regeneration, as well as delayed appearance of newly regenerated fibers. Recovery of wild-type BDNF levels was sufficient to restore normal regeneration. Together, these findings suggest that BDNF plays an important role in regulating satellite cell function and regeneration in vivo, particularly during early stages.
Figures
References
-
- Capsoni S., Ruberti F., Di Daniel E., Cattaneo A. Muscular dystrophy in adult and aged anti-NGF transgenic mice resembles an inclusion body myopathy. J. Neurosci. Res. 2000;59:553–560. - PubMed
-
- Carrasco D. I., English A. W. Neurotrophin 4/5 is required for the normal development of the slow muscle fiber phenotype in the rat soleus. J. Exp. Biol. 2003;206:2191–2200. - PubMed
-
- Charge S. B., Rudnicki M. A. Cellular and molecular regulation of muscle regeneration. Physiol. Rev. 2004;84:209–238. - PubMed
-
- Chevrel G., Hohlfeld R., Sendtner M. The role of neurotrophins in muscle under physiological and pathophysiological conditions. Muscle Nerve. 2006;33:462–476. - PubMed
-
- Condrea E. Membrane-active polypeptides from snake venom: cardiotoxins and haemocytotoxins. Experimentia. 1974;30:121–129. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
