

Spatial and temporal processing of single auditory cortical neurons and populations of neurons in the macaque monkey
- PMID: 20430079
- PMCID: PMC2941552
- DOI: 10.1016/j.heares.2010.03.084
Spatial and temporal processing of single auditory cortical neurons and populations of neurons in the macaque monkey
Abstract
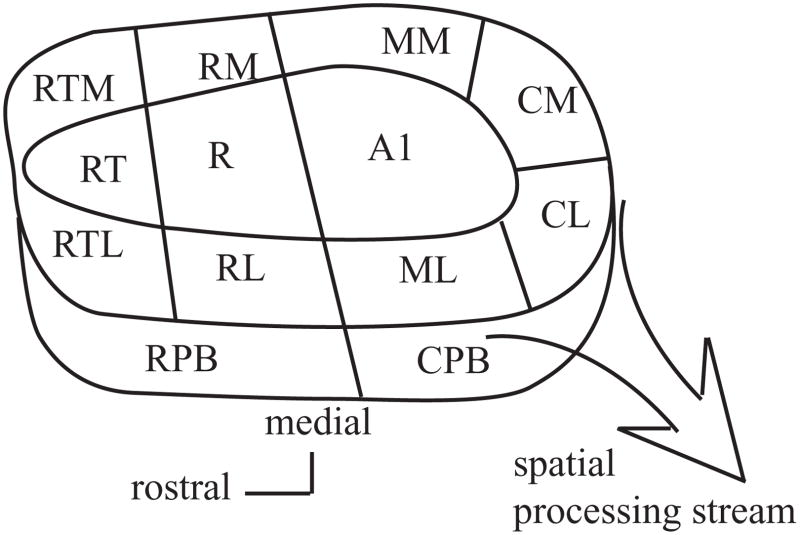
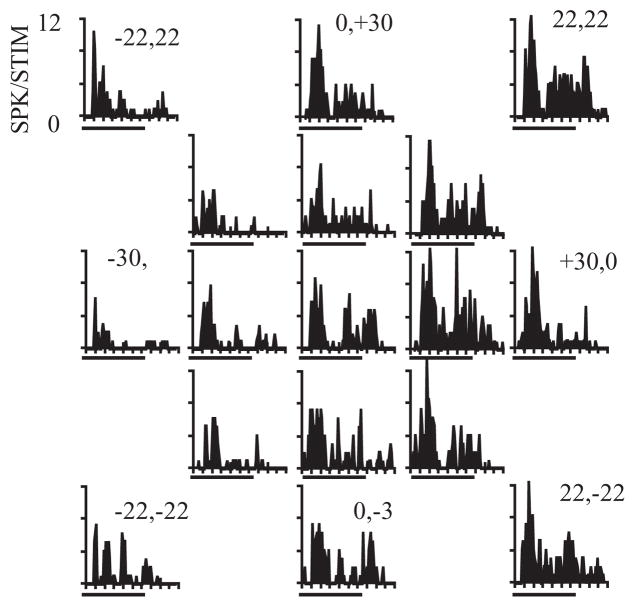
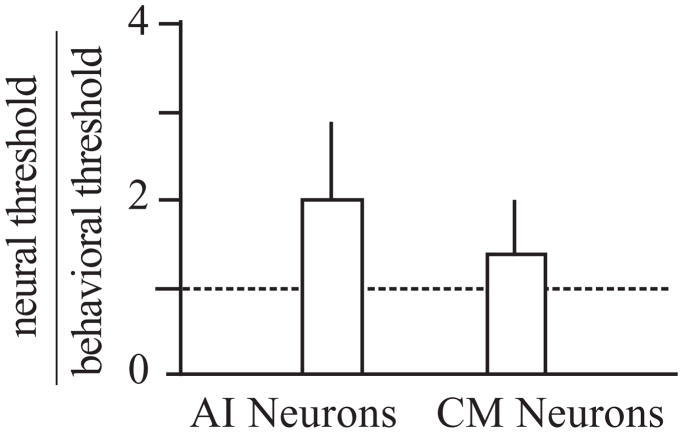
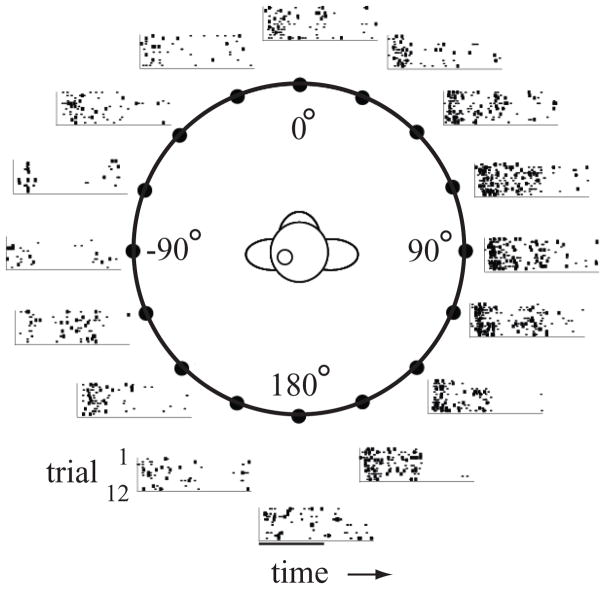
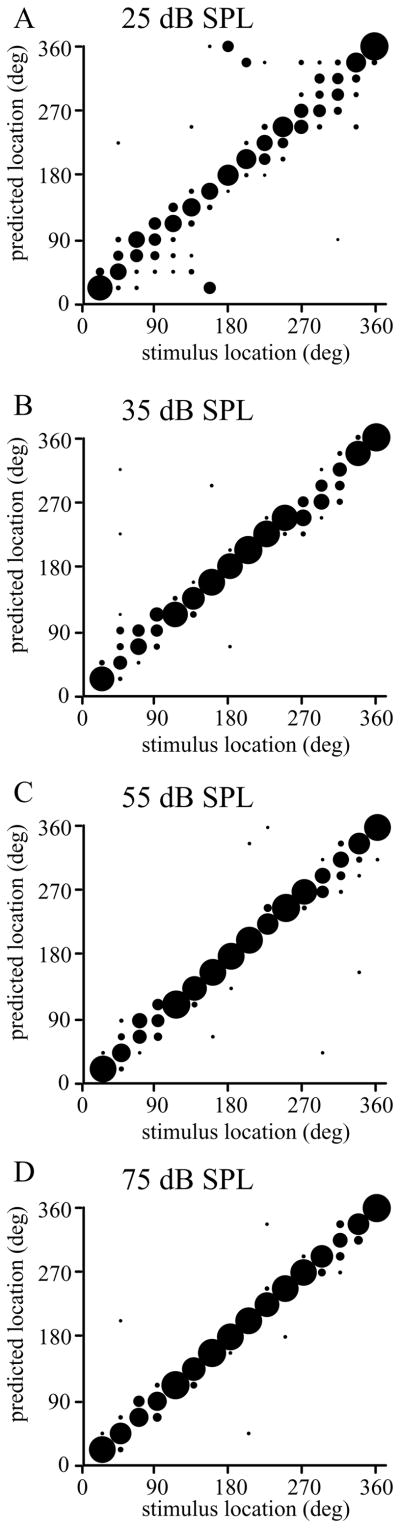
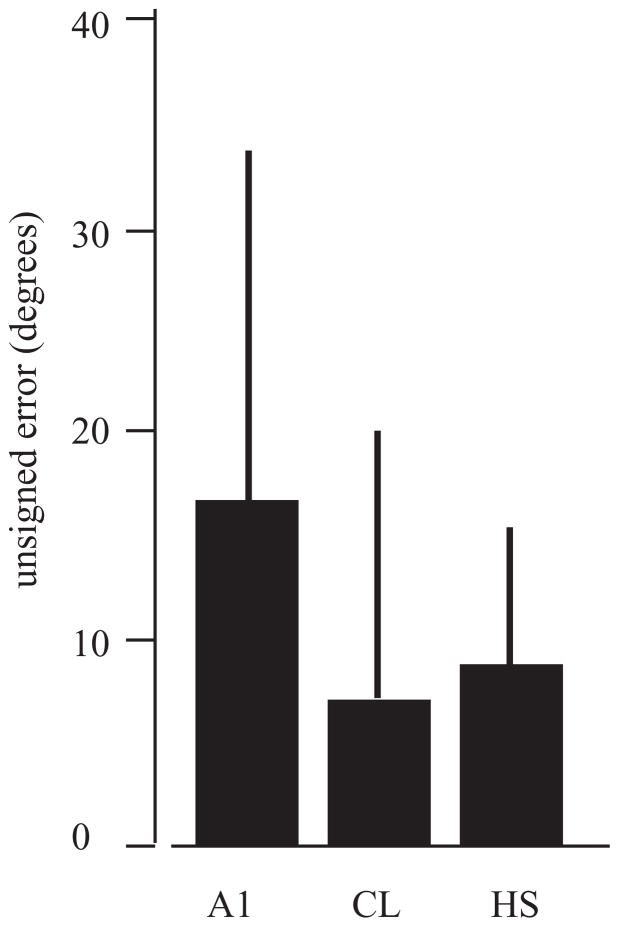
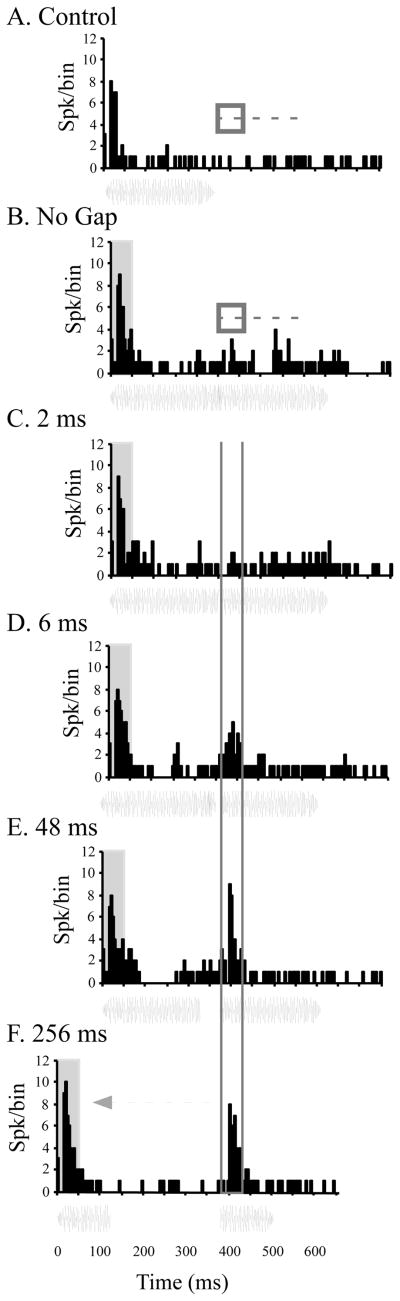
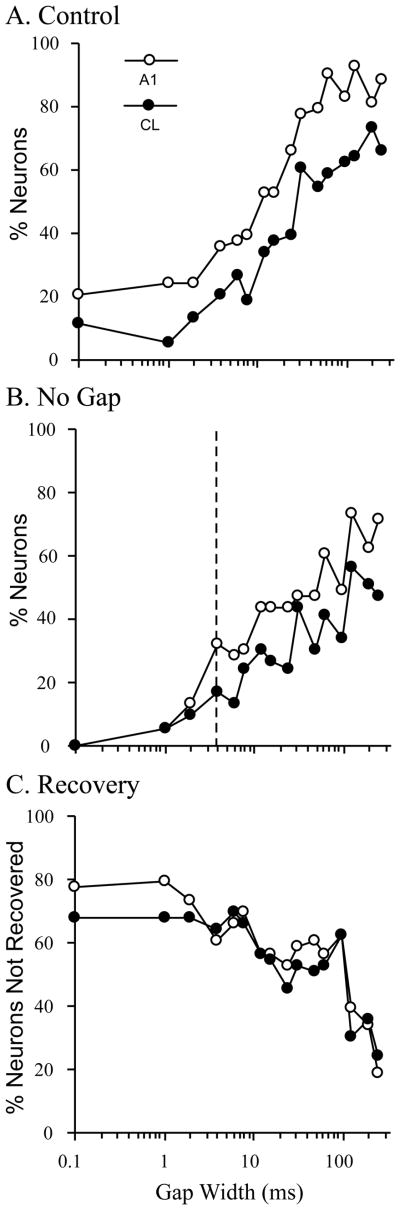
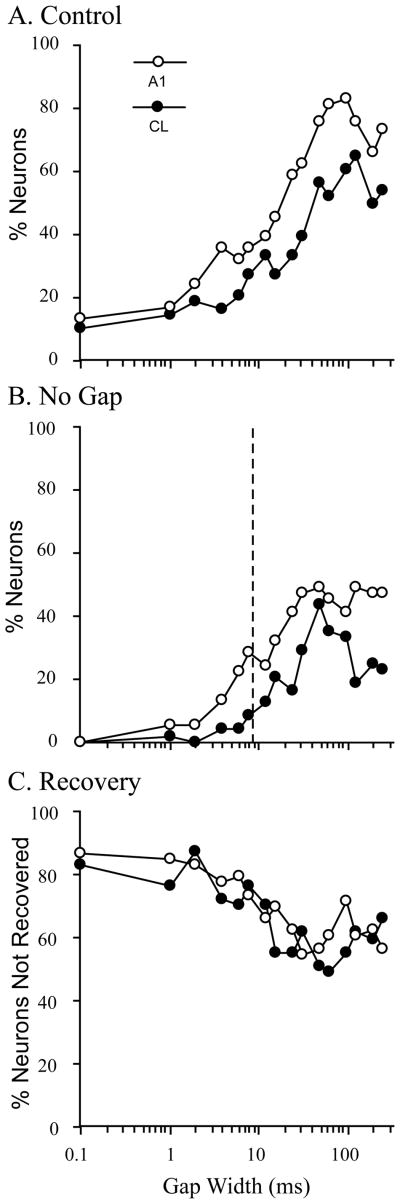
The auditory cortex is known to be a necessary neural structure for the perception of acoustic signals, particularly the spatial location and the temporal features of complex auditory stimuli. Previous studies have indicated that there is no topographic map of acoustic space in the auditory cortex and it has been proposed that spatial locations are represented by some sort of population code. Additionally, in spite of temporal processing deficits being one of the hallmark consequences of normal aging, the temporal coding of acoustic stimuli remains poorly understood. This report will address these two issues by discussing the results from several studies describing responses of single auditory cortical neurons in the non-human primate. First, we will review studies that have addressed potential spike-rate population codes of acoustic space in the caudal belt of auditory cortex. Second, we will present new data on the neuronal responses to gap stimuli in aged monkeys and compare them to published reports of gap detection thresholds. Together these studies indicate that the alert macaque monkey is an excellent model system to study both spatial and temporal processing in the auditory cortex at the single neuron level.
© 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Altshuler MW, Comalli PE. Effect of stimulus intensity and frequency on median horizontal plane sound localization. J Auditory Res. 1975;15:262–265.
-
- Beitel RE, Kaas JH. Effects of bilateral and unilateral ablation of auditory cortex in cats on the unconditioned head orienting response to acoustic stimuli. J Neurophysiol. 1993;70:351–369. - PubMed
-
- Bennett CL, Davis RT, Miller JM. Demonstration of presbycusis across repeated measures in a nonhuman primate species. Behav Neurosci. 1983;97:602–607. - PubMed
-
- Blauert J. The Psychophysics of Human Sound Localization. 2. MIT Press; 1997. Spatial Hearing.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
