

The acetyltransferase activity of the bacterial toxin YopJ of Yersinia is activated by eukaryotic host cell inositol hexakisphosphate
- PMID: 20430892
- PMCID: PMC2888404
- DOI: 10.1074/jbc.M110.126581
The acetyltransferase activity of the bacterial toxin YopJ of Yersinia is activated by eukaryotic host cell inositol hexakisphosphate
Abstract
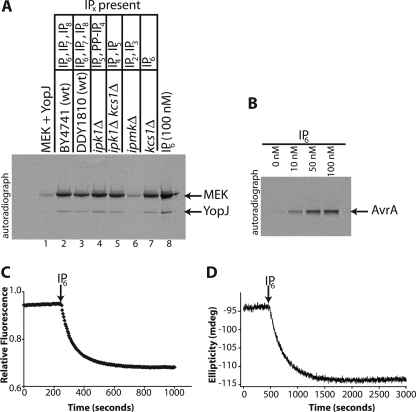
Plague, one of the most devastating diseases in human history, is caused by the bacterium Yersinia pestis. The bacteria use a syringe-like macromolecular assembly to secrete various toxins directly into the host cells they infect. One such Yersinia outer protein, YopJ, performs the task of dampening innate immune responses in the host by simultaneously inhibiting the MAPK and NFkappaB signaling pathways. YopJ catalyzes the transfer of acetyl groups to serine, threonine, and lysine residues on target proteins. Acetylation of serine and threonine residues prevents them from being phosphorylated thereby preventing the activation of signaling molecules on which they are located. In this study, we describe the requirement of a host-cell factor for full activation of the acetyltransferase activity of YopJ and identify this activating factor to be inositol hexakisphosphate (IP(6)). We extend the applicability of our results to show that IP(6) also stimulates the acetyltransferase activity of AvrA, the YopJ homologue from Salmonella typhimurium. Furthermore, an IP(6)-induced conformational change in AvrA suggests that IP(6) acts as an allosteric activator of enzyme activity. Our results suggest that YopJ-family enzymes are quiescent in the bacterium where they are synthesized, because bacteria lack IP(6); once injected into mammalian cells by the pathogen these toxins bind host cell IP(6), are activated, and deregulate the MAPK and NFkappaB signaling pathways thereby subverting innate immunity.
Figures
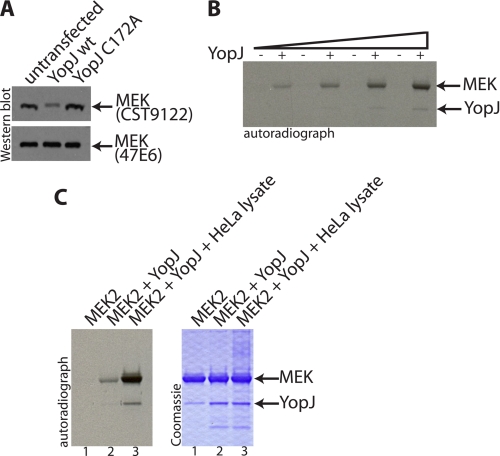
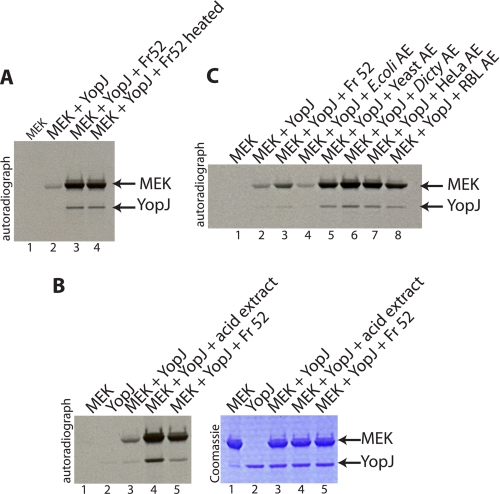
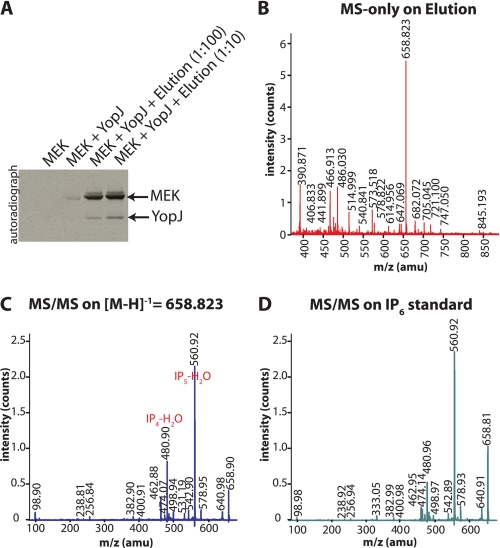
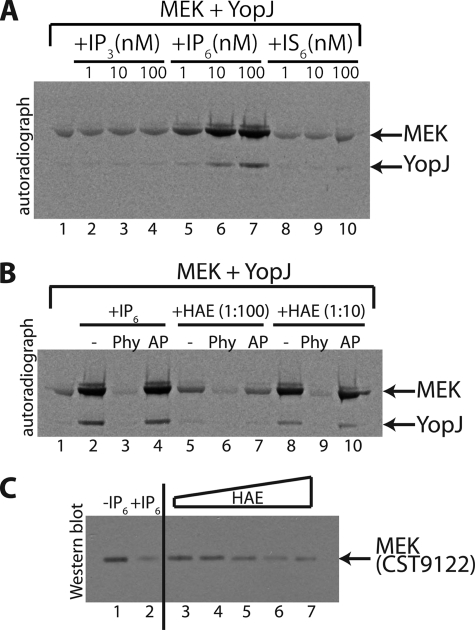
References
-
- Mueller C. A., Broz P., Cornelis G. R. (2008) Mol. Microbiol. 68, 1085–1095 - PubMed
-
- Trosky J. E., Liverman A. D., Orth K. (2008) Cell Microbiol. 10, 557–565 - PubMed
-
- Palmer L. E., Hobbie S., Galán J. E., Bliska J. B. (1998) Mol. Microbiol. 27, 953–965 - PubMed
-
- Mukherjee S., Keitany G., Li Y., Wang Y., Ball H. L., Goldsmith E. J., Orth K. (2006) Science 312, 1211–1214 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
