

Endothelial nitric oxide synthase deficiency causes collateral vessel rarefaction and impairs activation of a cell cycle gene network during arteriogenesis
- PMID: 20431061
- PMCID: PMC3401938
- DOI: 10.1161/CIRCRESAHA.109.212746
Endothelial nitric oxide synthase deficiency causes collateral vessel rarefaction and impairs activation of a cell cycle gene network during arteriogenesis
Abstract
Rationale: The collateral circulation is tissue- and life-saving in obstructive arterial disease. Disappointing outcomes in clinical trials aimed at augmenting collateral growth highlight the need for greater understanding of collateral biology.
Objective: The role of endothelial nitric oxide synthase (eNOS) in forming native (preexisting) collaterals and remodeling in obstructive disease are unknown or controversial issues, respectively.
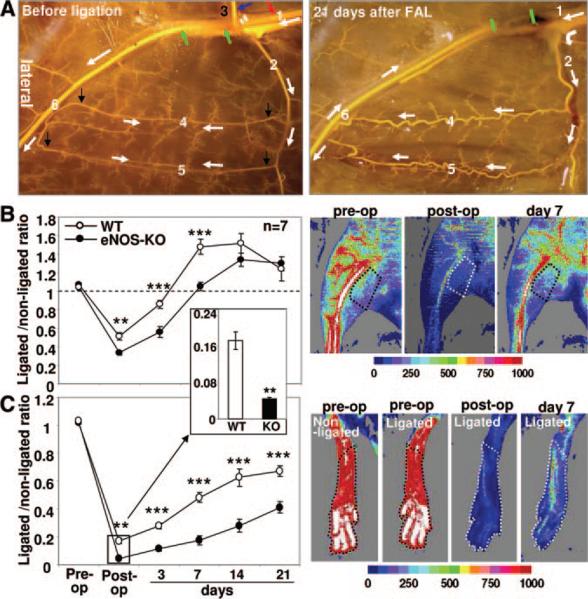
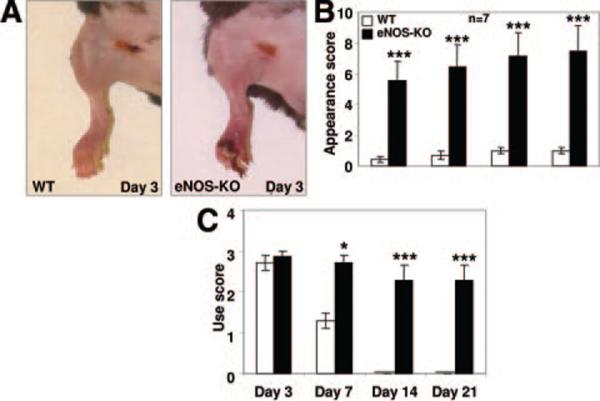
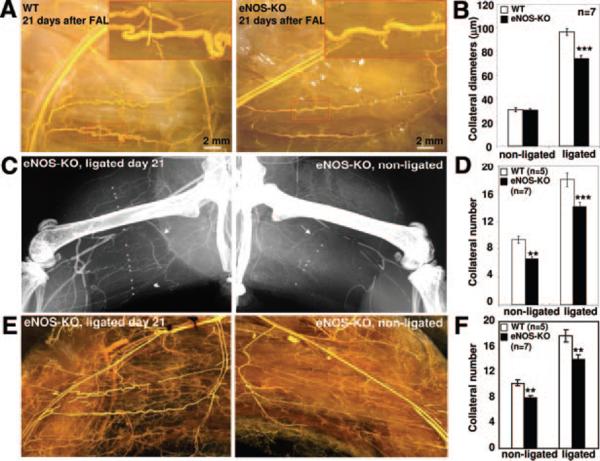
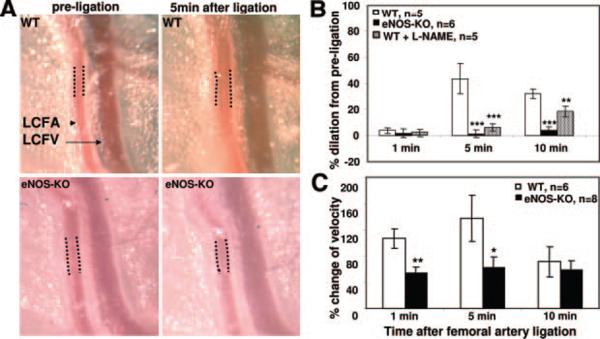
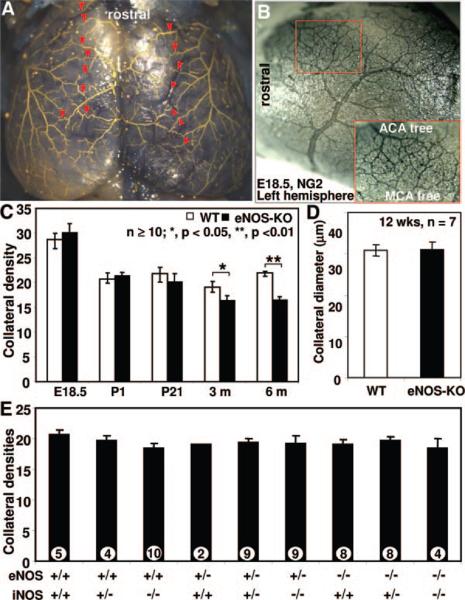
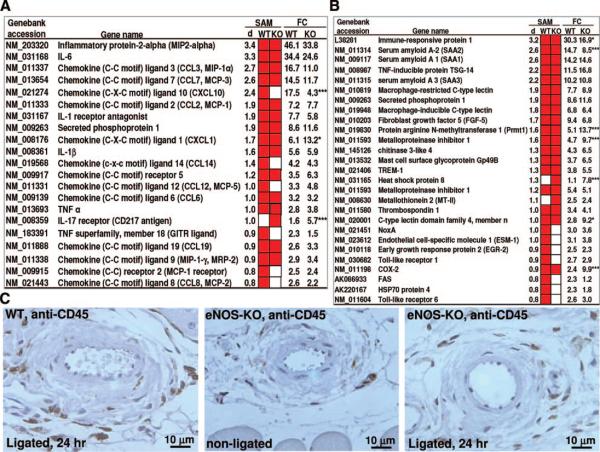
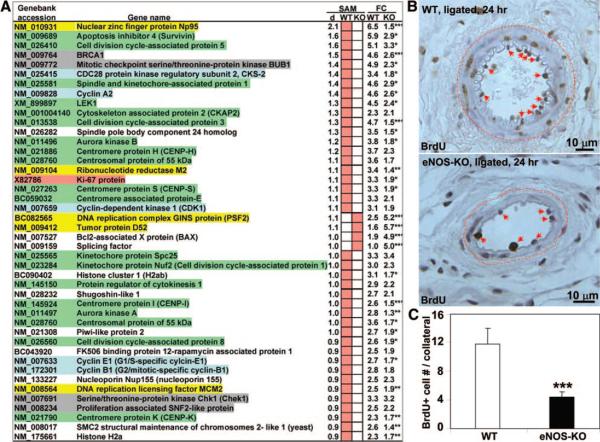
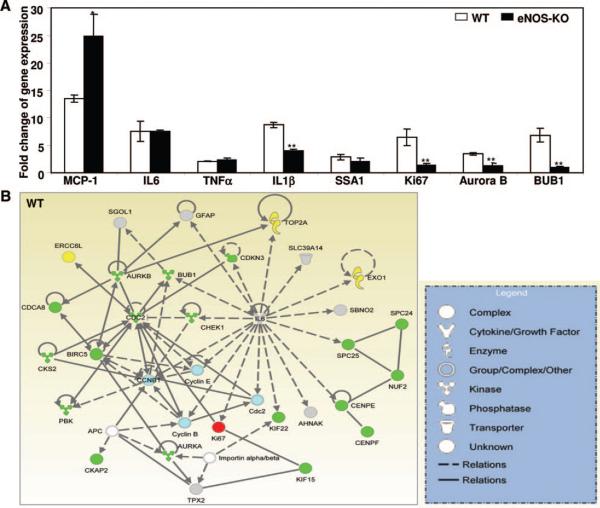
Methods and results: We compared the native collateral circulation in healthy tissue and collateral remodeling after femoral artery ligation (FAL) in wild-type and eNOS-knockout (KO) mice. Perfusion after FAL fell further in adult eNOS-KOs, in association with fewer native collaterals in hindlimb (confirmed in brain). This was not attributable to impaired collateral formation in the embryo-neonate, but rather from collateral loss during growth to adulthood. Compared to wild-type, eNOS-KOs evidenced reduced collateral remodeling, angiogenesis, and flow-mediated dilation of the arterial bed supplying the collaterals, resulting in lower perfusion and greater ischemic injury at all time points over 21 days following FAL. To probe the mechanism for impaired remodeling, we performed genome-wide expression profiling of isolated, remodeling hindlimb collaterals 24 hour after FAL. Upregulation of genes encoding cytokines/chemokines, inflammatory, stress response, and cell cycle proteins was evident in wild-type mice. In contrast, expression was lower in 40 of 44 cell cycle genes in eNOS-KO mice, in association with impaired proliferation of vascular wall cells.
Conclusions: Our findings suggest a novel role for eNOS in maintaining native collateral density during natural growth to adulthood and in collateral remodeling in obstructive disease, the latter through regulation of cell proliferation.
Figures
Similar articles
-
Mechanisms of Amplified Arteriogenesis in Collateral Artery Segments Exposed to Reversed Flow Direction.Arterioscler Thromb Vasc Biol. 2015 Nov;35(11):2354-65. doi: 10.1161/ATVBAHA.115.305775. Epub 2015 Sep 3. Arterioscler Thromb Vasc Biol. 2015. PMID: 26338297 Free PMC article.
-
Endothelial Hyaluronan Synthase 3 Augments Postischemic Arteriogenesis Through CD44/eNOS Signaling.Arterioscler Thromb Vasc Biol. 2021 Oct;41(10):2551-2562. doi: 10.1161/ATVBAHA.121.315478. Epub 2021 Aug 12. Arterioscler Thromb Vasc Biol. 2021. PMID: 34380333
-
P2Y2 nucleotide receptor mediates arteriogenesis in a murine model of hind limb ischemia.J Vasc Surg. 2016 Jan;63(1):216-25. doi: 10.1016/j.jvs.2014.06.112. Epub 2014 Jul 31. J Vasc Surg. 2016. PMID: 25088742 Free PMC article.
-
Arteriogenesis: the development and growth of collateral arteries.Microcirculation. 2003 Jan;10(1):83-97. doi: 10.1038/sj.mn.7800173. Microcirculation. 2003. PMID: 12610665 Review.
-
Influence of inflammatory cytokines on arteriogenesis.Microcirculation. 2003 Jun;10(3-4):371-9. doi: 10.1038/sj.mn.7800199. Microcirculation. 2003. PMID: 12851653 Review.
Cited by
-
Minicircle DNA-mediated endothelial nitric oxide synthase gene transfer enhances angiogenic responses of bone marrow-derived mesenchymal stem cells.Stem Cell Res Ther. 2016 Apr 1;7:48. doi: 10.1186/s13287-016-0307-2. Stem Cell Res Ther. 2016. PMID: 27036881 Free PMC article.
-
Neuroprotective Effects of Simvastatin and Cilostazol in L-Methionine-Induced Vascular Dementia in Rats.Mol Neurobiol. 2017 Sep;54(7):5074-5084. doi: 10.1007/s12035-016-0051-8. Epub 2016 Aug 20. Mol Neurobiol. 2017. PMID: 27544235
-
Nox2 and p47(phox) modulate compensatory growth of primary collateral arteries.Am J Physiol Heart Circ Physiol. 2014 May 15;306(10):H1435-43. doi: 10.1152/ajpheart.00828.2013. Epub 2014 Mar 14. Am J Physiol Heart Circ Physiol. 2014. PMID: 24633549 Free PMC article.
-
RGS5 promotes arterial growth during arteriogenesis.EMBO Mol Med. 2014 Aug;6(8):1075-89. doi: 10.15252/emmm.201403864. EMBO Mol Med. 2014. PMID: 24972930 Free PMC article.
-
Therapeutic ultrasound reverses peripheral ischemia in type 2 diabetic mice through PI3K-Akt-eNOS pathway.Am J Transl Res. 2016 Sep 15;8(9):3666-3677. eCollection 2016. Am J Transl Res. 2016. PMID: 27725849 Free PMC article.
References
-
- Meier P, Gloekler S, Zbinden R, Beckh S, de Marchi SF, Zbinden S, Wustmann K, Billinger M, Vogel R, Cook S, Wenaweser P, Togni M, Windecker S, Meier B, Seiler C. Beneficial effect of recruitable collaterals: a 10-year follow-up study in patients with stable coronary artery disease undergoing quantitative collateral measurements. Circulation. 2007;116:975–983. - PubMed
-
- Chalothorn D, Clayton JA, Zhang H, Pomp D, Faber JE. Collateral density, remodeling, and VEGF-A expression differ widely between mouse strains. Physiol Genomics. 2007;30:179–191. - PubMed
-
- Chalothorn D, Faber JE. Differences in collateral formation in the embryo are associated with genetic variation in patterning and maturation of the cerebral cortical circulation. Arterioscler Thromb Vasc Biol. 2009;29:250. Abstract.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
