

The Puf RNA-binding proteins FBF-1 and FBF-2 inhibit the expression of synaptonemal complex proteins in germline stem cells
- PMID: 20431119
- PMCID: PMC2867315
- DOI: 10.1242/dev.050799
The Puf RNA-binding proteins FBF-1 and FBF-2 inhibit the expression of synaptonemal complex proteins in germline stem cells
Abstract
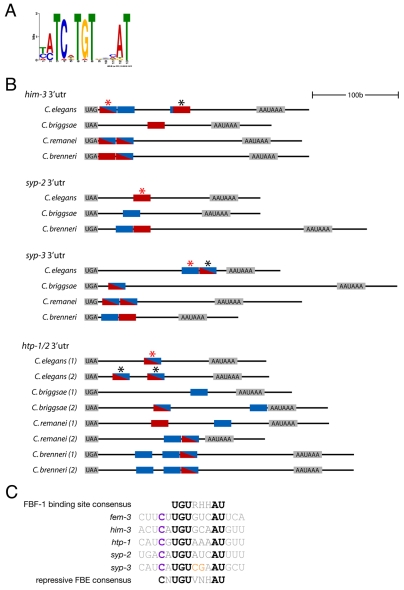
FBF-1 and FBF-2 (collectively FBF) are two nearly identical Puf-domain RNA-binding proteins that regulate the switch from mitosis to meiosis in the C. elegans germline. In germline stem cells, FBF prevents premature meiotic entry by inhibiting the expression of meiotic regulators, such as the RNA-binding protein GLD-1. Here, we demonstrate that FBF also directly inhibits the expression of structural components of meiotic chromosomes. HIM-3, HTP-1, HTP-2, SYP-2 and SYP-3 are components of the synaptonemal complex (SC) that forms between homologous chromosomes during meiotic prophase. In wild-type germlines, the five SC proteins are expressed shortly before meiotic entry. This pattern depends on FBF binding sites in the 3' UTRs of the SC mRNAs. In the absence of FBF or the FBF binding sites, SC proteins are expressed precociously in germline stem cells and their precursors. SC proteins aggregate and SC formation fails at meiotic entry. Precocious SC protein expression is observed even when meiotic entry is delayed in fbf mutants by reducing GLD-1. We propose that parallel regulation by FBF ensures that in wild-type gonads, meiotic entry is coordinated with just-in-time synthesis of synaptonemal proteins.
Figures





Similar articles
-
A conserved RNA-binding protein controls germline stem cells in Caenorhabditis elegans.Nature. 2002 Jun 6;417(6889):660-3. doi: 10.1038/nature754. Epub 2002 May 22. Nature. 2002. PMID: 12050669
-
C. elegans RNA-binding proteins PUF-8 and MEX-3 function redundantly to promote germline stem cell mitosis.Dev Biol. 2009 Feb 15;326(2):295-304. doi: 10.1016/j.ydbio.2008.11.024. Epub 2008 Dec 7. Dev Biol. 2009. PMID: 19100255 Free PMC article.
-
Cyclin E and Cdk2 control GLD-1, the mitosis/meiosis decision, and germline stem cells in Caenorhabditis elegans.PLoS Genet. 2011 Mar;7(3):e1001348. doi: 10.1371/journal.pgen.1001348. Epub 2011 Mar 24. PLoS Genet. 2011. PMID: 21455289 Free PMC article.
-
The regulatory network controlling the proliferation-meiotic entry decision in the Caenorhabditis elegans germ line.Curr Top Dev Biol. 2006;76:185-215. doi: 10.1016/S0070-2153(06)76006-9. Curr Top Dev Biol. 2006. PMID: 17118267 Review.
-
Role of PUF-8/PUF protein in stem cell control, sperm-oocyte decision and cell fate reprogramming.J Cell Physiol. 2014 Oct;229(10):1306-11. doi: 10.1002/jcp.24618. J Cell Physiol. 2014. PMID: 24638209 Review.
Cited by
-
CRL2(LRR-1) E3-ligase regulates proliferation and progression through meiosis in the Caenorhabditis elegans germline.PLoS Genet. 2013 Mar;9(3):e1003375. doi: 10.1371/journal.pgen.1003375. Epub 2013 Mar 28. PLoS Genet. 2013. PMID: 23555289 Free PMC article.
-
Role of the CRL2(LRR-1) E3 ubiquitin-ligase in the development of the germline in C. elegans.Worm. 2013 Jul 1;2(3):e25716. doi: 10.4161/worm.25716. Epub 2013 Jul 17. Worm. 2013. PMID: 24778939 Free PMC article.
-
Germline Stem Cell Differentiation Entails Regional Control of Cell Fate Regulator GLD-1 in Caenorhabditis elegans.Genetics. 2016 Mar;202(3):1085-103. doi: 10.1534/genetics.115.185678. Epub 2016 Jan 12. Genetics. 2016. PMID: 26757772 Free PMC article.
-
A pumilio homolog in Polycelis sp.Dev Genes Evol. 2014 Feb;224(1):53-6. doi: 10.1007/s00427-013-0459-3. Epub 2013 Nov 30. Dev Genes Evol. 2014. PMID: 24292205
-
PUF-8 suppresses the somatic transcription factor PAL-1 expression in C. elegans germline stem cells.Dev Biol. 2011 Dec 1;360(1):195-207. doi: 10.1016/j.ydbio.2011.09.021. Epub 2011 Sep 24. Dev Biol. 2011. PMID: 21968099 Free PMC article.
References
-
- Ahringer J., Kimble J. (1991). Control of the sperm-oocyte switch in Caenorhabditis elegans hermaphrodites by the fem-3 3′ untranslated region. Nature 349, 346-348 - PubMed
-
- Bailey T. L., Elkan C. (1994). Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 2, 28-36 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
