

PAR-3 mediates the initial clustering and apical localization of junction and polarity proteins during C. elegans intestinal epithelial cell polarization
- PMID: 20431121
- PMCID: PMC2867319
- DOI: 10.1242/dev.047647
PAR-3 mediates the initial clustering and apical localization of junction and polarity proteins during C. elegans intestinal epithelial cell polarization
Abstract
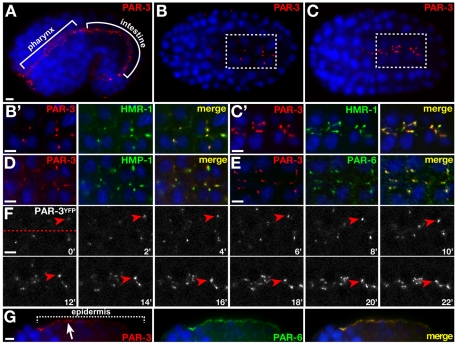
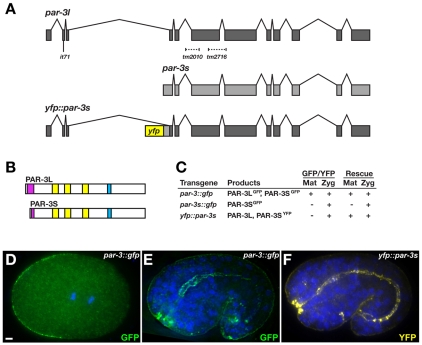
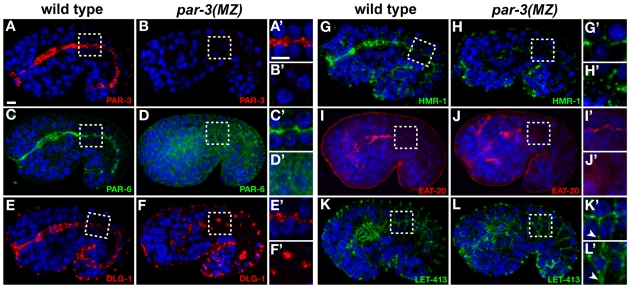
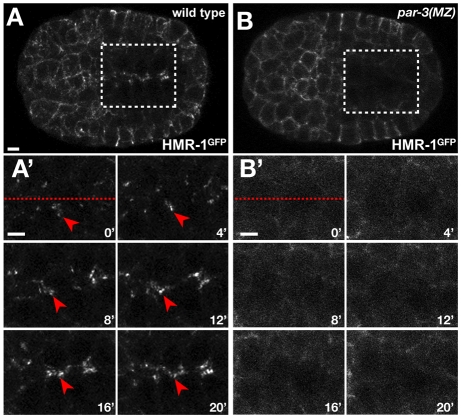
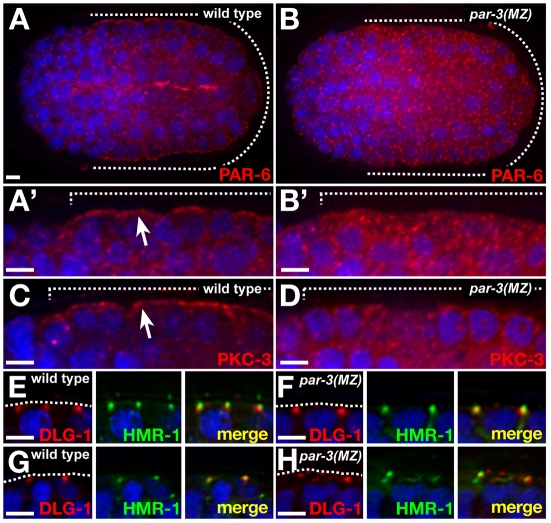
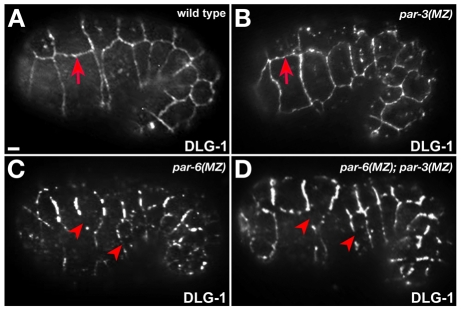
The apicobasal polarity of epithelial cells is critical for organ morphogenesis and function, and loss of polarity can promote tumorigenesis. Most epithelial cells form when precursor cells receive a polarization cue, develop distinct apical and basolateral domains and assemble junctions near their apical surface. The scaffolding protein PAR-3 regulates epithelial cell polarity, but its cellular role in the transition from precursor cell to polarized epithelial cell has not been determined in vivo. Here, we use a targeted protein-degradation strategy to remove PAR-3 from C. elegans embryos and examine its cellular role as intestinal precursor cells become polarized epithelial cells. At initial stages of polarization, PAR-3 accumulates in cortical foci that contain E-cadherin, other adherens junction proteins, and the polarity proteins PAR-6 and PKC-3. Using live imaging, we show that PAR-3 foci move apically and cluster, and that PAR-3 is required to assemble E-cadherin into foci and for foci to accumulate at the apical surface. We propose that PAR-3 facilitates polarization by promoting the initial clustering of junction and polarity proteins that then travel and accumulate apically. Unexpectedly, superficial epidermal cells form apical junctions in the absence of PAR-3, and we show that PAR-6 has a PAR-3-independent role in these cells to promote apical junction maturation. These findings indicate that PAR-3 and PAR-6 function sequentially to position and mature apical junctions, and that the requirement for PAR-3 can vary in different types of epithelial cells.
Figures

References
-
- Aono S., Legouis R., Hoose W. A., Kemphues K. J. (2004). PAR-3 is required for epithelial cell polarity in the distal spermatheca of C. elegans. Development 131, 2865-2874 - PubMed
-
- Bilder D. (2004). Epithelial polarity and proliferation control: links from the Drosophila neoplastic tumor suppressors. Genes Dev. 18, 1909-1925 - PubMed
-
- Bossinger O., Klebes A., Segbert C., Theres C., Knust E. (2001). Zonula adherens formation in Caenorhabditis elegans requires dlg-1, the homologue of the Drosophila gene discs large. Dev. Biol. 230, 29-42 - PubMed
-
- Bossinger O., Fukushige T., Claeys M., Borgonie G., McGhee J. D. (2004). The apical disposition of the Caenorhabditis elegans intestinal terminal web is maintained by LET-413. Dev. Biol. 268, 448-456 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
