

Endocytic and secretory traffic in Arabidopsis merge in the trans-Golgi network/early endosome, an independent and highly dynamic organelle
- PMID: 20435907
- PMCID: PMC2879741
- DOI: 10.1105/tpc.109.072637
Endocytic and secretory traffic in Arabidopsis merge in the trans-Golgi network/early endosome, an independent and highly dynamic organelle
Abstract
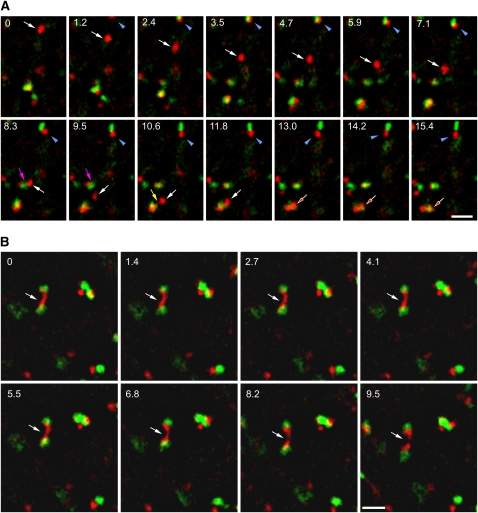
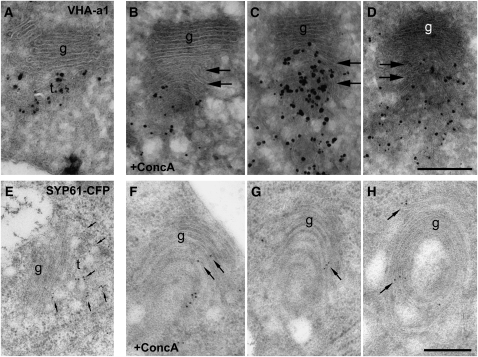
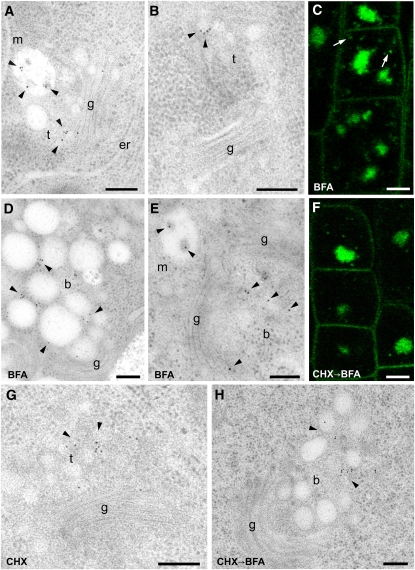
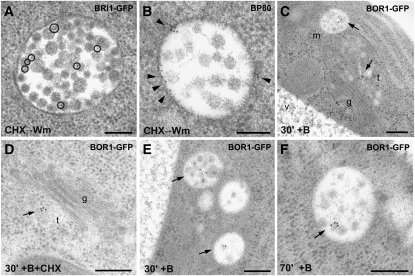
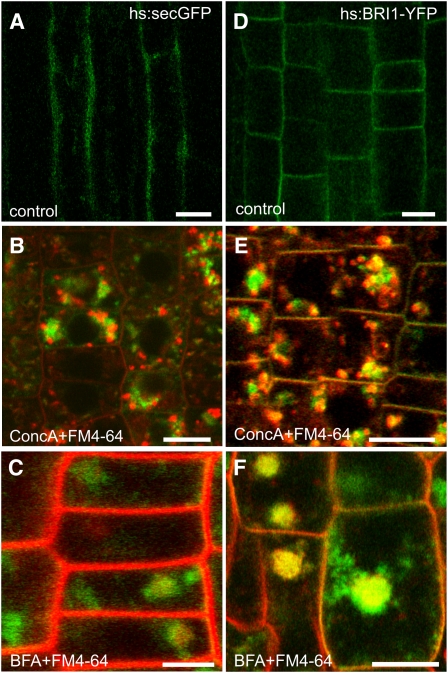
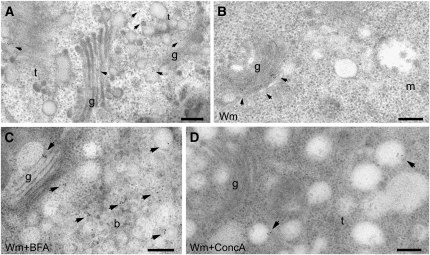
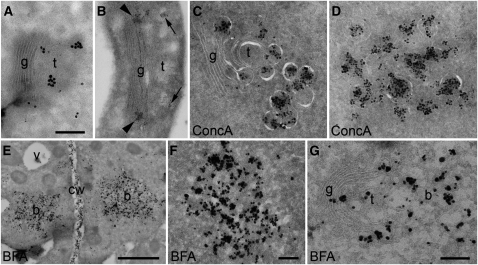
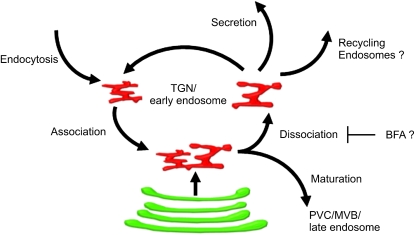
Plants constantly adjust their repertoire of plasma membrane proteins that mediates transduction of environmental and developmental signals as well as transport of ions, nutrients, and hormones. The importance of regulated secretory and endocytic trafficking is becoming increasingly clear; however, our knowledge of the compartments and molecular machinery involved is still fragmentary. We used immunogold electron microscopy and confocal laser scanning microscopy to trace the route of cargo molecules, including the BRASSINOSTEROID INSENSITIVE1 receptor and the REQUIRES HIGH BORON1 boron exporter, throughout the plant endomembrane system. Our results provide evidence that both endocytic and secretory cargo pass through the trans-Golgi network/early endosome (TGN/EE) and demonstrate that cargo in late endosomes/multivesicular bodies is destined for vacuolar degradation. Moreover, using spinning disc microscopy, we show that TGN/EEs move independently and are only transiently associated with an individual Golgi stack.
Figures
References
-
- Baluska F., Liners F., Hlavacka A., Schlicht M., Van Cutsem P., McCurdy D.W., Menzel D. (2005). Cell wall pectins and xyloglucans are internalized into dividing root cells and accumulate within cell plates during cytokinesis. Protoplasma 225: 141–155 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
