

The nuclear export receptor XPO-1 supports primary miRNA processing in C. elegans and Drosophila
- PMID: 20436454
- PMCID: PMC2885935
- DOI: 10.1038/emboj.2010.82
The nuclear export receptor XPO-1 supports primary miRNA processing in C. elegans and Drosophila
Abstract
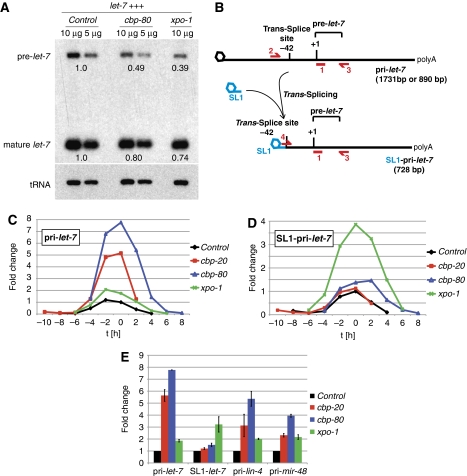
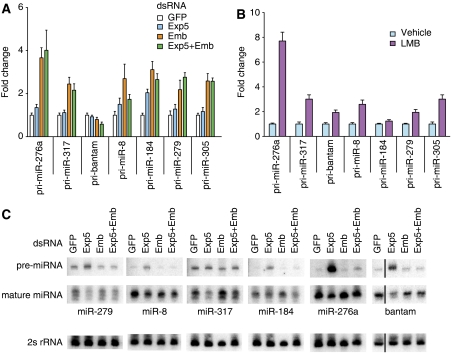
MicroRNA (miRNA) biogenesis proceeds from a primary transcript (pri-miRNA) through the pre-miRNA into the mature miRNA. Here, we identify a role of the Caenorhabditis elegans nuclear export receptor XPO-1 and the cap-binding proteins CBP-20/NCBP-2 and CBP-80/NCBP-1 in this process. The RNA-mediated interference of any of these genes causes retarded heterochronic phenotypes similar to those observed for animals with mutations in the let-7 miRNA or core miRNA machinery genes. Moreover, pre- and mature miRNAs become depleted, whereas primary miRNA transcripts accumulate. An involvement of XPO-1 in miRNA biogenesis is conserved in Drosophila, in which knockdown of Embargoed/XPO-1 or its chemical inhibition through leptomycin B causes pri-miRNA accumulation. Our findings demonstrate that XPO-1/Emb promotes the pri-miRNA-to-pre-miRNA processing and we propose that this function involves intranuclear transport and/or nuclear export of primary miRNAs.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Boulon S, Verheggen C, Jady BE, Girard C, Pescia C, Paul C, Ospina JK, Kiss T, Matera AG, Bordonné R, Bertrand E (2004) PHAX and CRM1 are required sequentially to transport U3 snoRNA to nucleoli. Mol Cell 16: 777–787 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
