

Cyclooxygenase-2 generates anti-inflammatory mediators from omega-3 fatty acids
- PMID: 20436486
- PMCID: PMC3077712
- DOI: 10.1038/nchembio.367
Cyclooxygenase-2 generates anti-inflammatory mediators from omega-3 fatty acids
Abstract
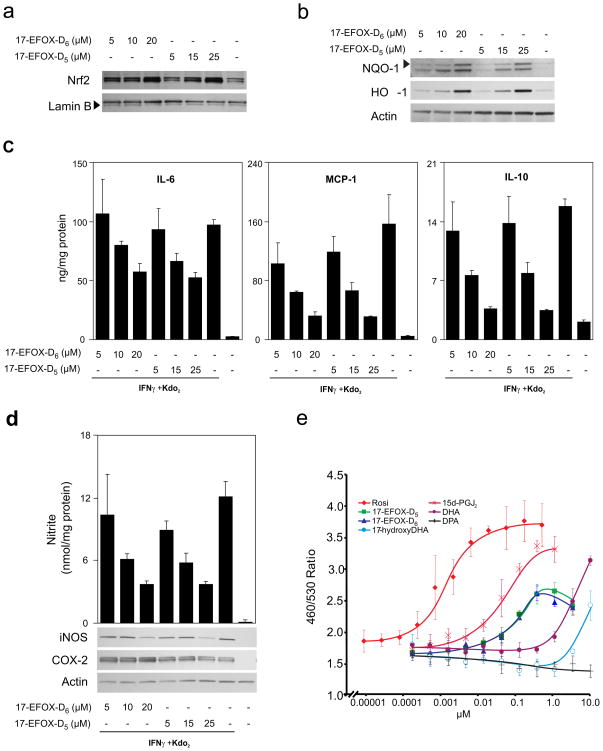
Electrophilic fatty acids are generated during inflammation by non-enzymatic reactions and can modulate inflammatory responses. We used a new mass spectrometry-based electrophile capture strategy to reveal the formation of electrophilic oxo-derivatives (EFOX) from the omega-3 fatty acids docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA). These EFOX were generated by a cyclooxygenase-2 (COX-2)-catalyzed mechanism in activated macrophages. Modulation of COX-2 activity by aspirin increased the rate of EFOX production and their intracellular levels. Owing to their electrophilic nature, EFOX adducted to cysteine and histidine residues of proteins and activated Nrf2-dependent anti-oxidant gene expression. We confirmed the anti-inflammatory nature of DHA- and DPA-derived EFOX by showing that they can act as peroxisome proliferator-activated receptor-gamma (PPAR gamma) agonists and inhibit pro-inflammatory cytokine and nitric oxide production, all within biological concentration ranges. These data support the idea that EFOX are signaling mediators that transduce the beneficial clinical effects of omega-3 fatty acids, COX-2 and aspirin.
Conflict of interest statement
Figures
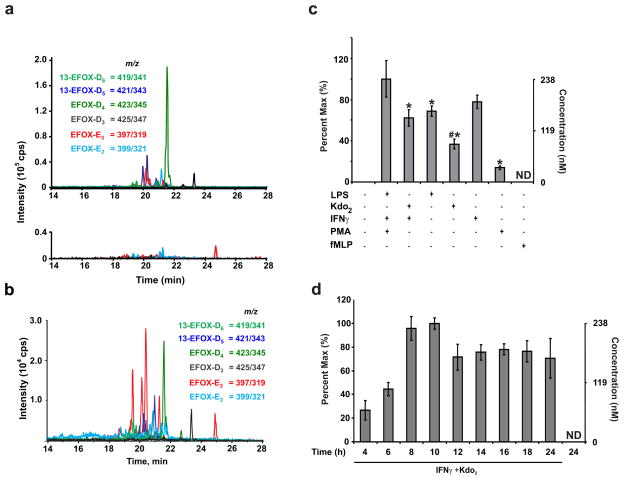
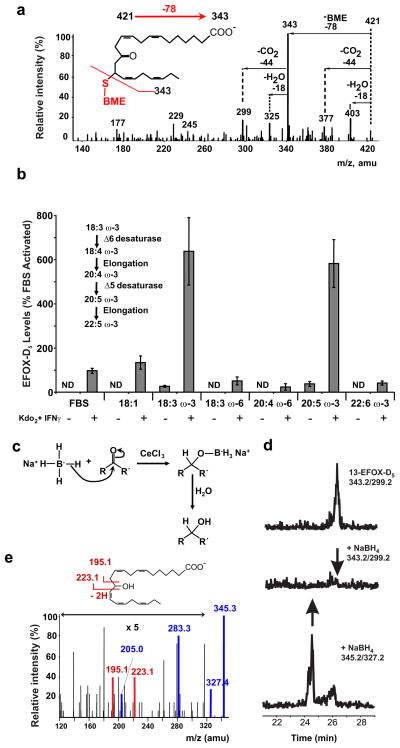
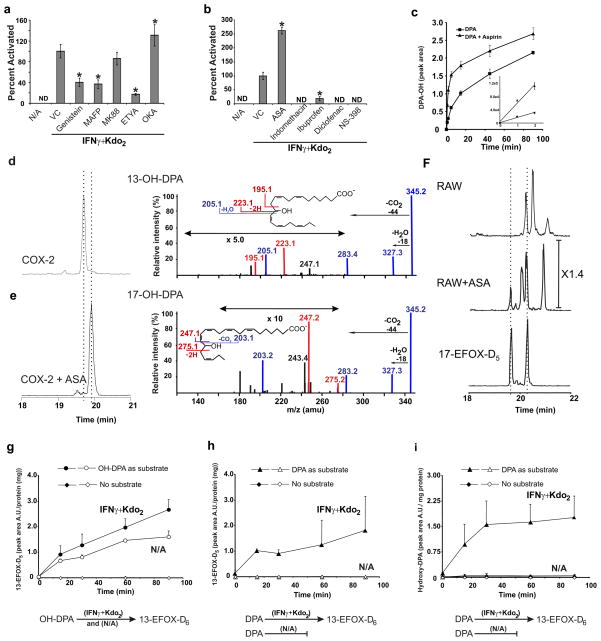
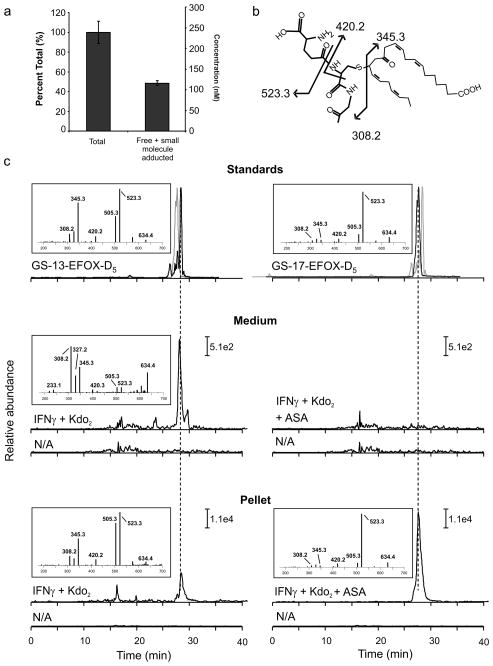
Comment in
-
COX-2's new role in inflammation.Nat Chem Biol. 2010 Jun;6(6):401-2. doi: 10.1038/nchembio.375. Nat Chem Biol. 2010. PMID: 20479749 No abstract available.
References
-
- Connor WE. Importance of n-3 fatty acids in health and disease. Am J Clin Nutr. 2000;71:171S–175S. - PubMed
-
- Neuringer M, Anderson GJ, Connor WE. The essentiality of n-3 fatty acids for the development and function of the retina and brain. Annu Rev Nutr. 1988;8:517–541. - PubMed
-
- Morris MC, Evans DA, Tangney CC, Bienias JL, Wilson RS. Fish consumption and cognitive decline with age in a large community study. Arch Neurol. 2005;62:1849–1853. - PubMed
-
- Fedor D, Kelley DS. Prevention of insulin resistance by n-3 polyunsaturated fatty acids. Curr Opin Clin Nutr Metab Care. 2009;12:138–146. - PubMed
-
- Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin E after myocardial infarction: results of the GISSI-Prevenzione trial. Gruppo Italiano per lo Studio della Sopravvivenza nell’Infarto miocardico. Lancet. 1999;354:447–455. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
