

Repeated evolution of reproductive isolation in a marine snail: unveiling mechanisms of speciation
- PMID: 20439278
- PMCID: PMC2871885
- DOI: 10.1098/rstb.2009.0256
Repeated evolution of reproductive isolation in a marine snail: unveiling mechanisms of speciation
Abstract
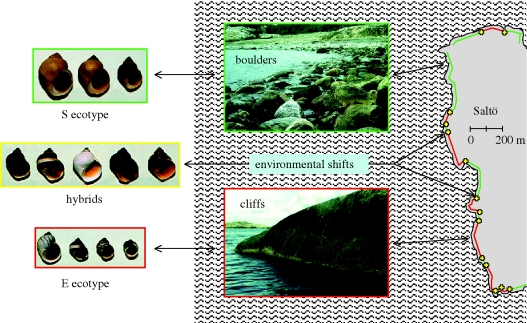
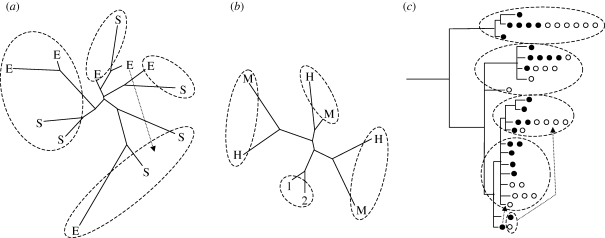
Distinct ecotypes of the snail Littorina saxatilis, each linked to a specific shore microhabitat, form a mosaic-like pattern with narrow hybrid zones in between, over which gene flow is 10-30% of within-ecotype gene flow. Multi-locus comparisons cluster populations by geographic affinity independent of ecotype, while loci under selection group populations by ecotype. The repeated occurrence of partially reproductively isolated ecotypes and the conflicting patterns in neutral and selected genes can either be explained by separation in allopatry followed by secondary overlap and extensive introgression that homogenizes neutral differences evolved under allopatry, or by repeated evolution in parapatry, or in sympatry, with the same ecotypes appearing in each local site. Data from Spain, the UK and Sweden give stronger support for a non-allopatric model of ecotype formation than for an allopatric model. Several different non-allopatric mechanisms can, however, explain the repeated evolution of the ecotypes: (i) parallel evolution by new mutations in different populations; (ii) evolution from standing genetic variation; and (iii) evolution in concert with rapid spread of new positive mutations among populations inhabiting similar environments. These models make different predictions that can be tested using comprehensive phylogenetic information combined with candidate loci sequencing.
Figures

Similar articles
-
The role of local ecology during hybridization at the initial stages of ecological speciation in a marine snail.J Evol Biol. 2013 Jul;26(7):1472-87. doi: 10.1111/jeb.12152. Epub 2013 May 11. J Evol Biol. 2013. PMID: 23663115
-
Case studies and mathematical models of ecological speciation. 3: Ecotype formation in a Swedish snail.Mol Ecol. 2009 Oct;18(19):4006-23. doi: 10.1111/j.1365-294X.2009.04320.x. Epub 2009 Sep 15. Mol Ecol. 2009. PMID: 19754510
-
Nonallopatric and parallel origin of local reproductive barriers between two snail ecotypes.Mol Ecol. 2004 Nov;13(11):3415-24. doi: 10.1111/j.1365-294X.2004.02330.x. Mol Ecol. 2004. PMID: 15488000
-
Review. Sympatric, parapatric or allopatric: the most important way to classify speciation?Philos Trans R Soc Lond B Biol Sci. 2008 Sep 27;363(1506):2997-3007. doi: 10.1098/rstb.2008.0076. Philos Trans R Soc Lond B Biol Sci. 2008. PMID: 18522915 Free PMC article. Review.
-
What can be learnt from a snail?Evol Appl. 2015 Jul 7;9(1):153-65. doi: 10.1111/eva.12277. eCollection 2016 Jan. Evol Appl. 2015. PMID: 27087845 Free PMC article. Review.
Cited by
-
Heterogeneous genome divergence, differential introgression, and the origin and structure of hybrid zones.Mol Ecol. 2016 Jun;25(11):2454-66. doi: 10.1111/mec.13582. Epub 2016 Mar 17. Mol Ecol. 2016. PMID: 26857437 Free PMC article.
-
Species and gene divergence in Littorina snails detected by array comparative genomic hybridization.BMC Genomics. 2014 Aug 18;15(1):687. doi: 10.1186/1471-2164-15-687. BMC Genomics. 2014. PMID: 25135785 Free PMC article.
-
Coupling of twelve putative chromosomal inversions maintains a strong barrier to gene flow between snail ecotypes.Evol Lett. 2024 Apr 23;8(4):575-586. doi: 10.1093/evlett/qrae014. eCollection 2024 Aug. Evol Lett. 2024. PMID: 39479507 Free PMC article.
-
Micro-spatial distribution of two sibling periwinkle species across the intertidal indicates hybrdization.Genetica. 2013 Sep;141(7-9):293-301. doi: 10.1007/s10709-013-9728-3. Epub 2013 Jul 26. Genetica. 2013. PMID: 23887893
-
Is embryo abortion a post-zygotic barrier to gene flow between Littorina ecotypes?J Evol Biol. 2020 Mar;33(3):342-351. doi: 10.1111/jeb.13570. Epub 2019 Dec 7. J Evol Biol. 2020. PMID: 31724256 Free PMC article.
References
-
- Andersen O., Wetten O. F., De Rosa M. C., Andre C., Alinovi C. C., Colafranceschi M., Brix O., Colosimo A.2009Haemoglobin polymorphisms affect the oxygen-binding properties in Atlantic cod populations. Proc. R. Soc. B 276, 833–841 (doi:10.1098/rspb.2008.1529) - DOI - PMC - PubMed
-
- Arendt J., Reznick D.2007Convergence and parallelism reconsidered: what have we learned about the genetics of adaptation? Trends Ecol. Evol. 23, 26–32 (doi:10.1016/j.tree.2007.09.011) - DOI - PubMed
-
- Barrett R. D. H., Schluter D.2007Adaptation from standing genetic variation. Trends Ecol. Evol. 23, 38–44 (doi:10.1016/j.tree.2007.09.008) - DOI - PubMed
-
- Bolnick D. I., Fitzpatrick B. M.2007Sympatric speciation: models and empirical evidence. Annu. Rev. Ecol. Evol. Syst. 38, 459–487 (doi:10.1146/annurev.ecolsys.38.091206.095804) - DOI
-
- Butlin R. K., Galindo J., Grahame J.2008Sympatric, parapatric or allopatric: the most important way to classify speciation? Phil. Trans. R. Soc. B 363, 2997–3007 (doi:10.1098/rstb.2008.0076) - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
