

doi: 10.1101/gad.1902110.
MVH in piRNA processing and gene silencing of retrotransposons
Affiliations
- PMID: 20439430
- PMCID: PMC2861188
- DOI: 10.1101/gad.1902110
Item in Clipboard
MVH in piRNA processing and gene silencing of retrotransposons
Genes Dev.
2010 May.
Abstract
VASA is an evolutionarily conserved RNA helicase essential for germ cell development. The mouse PIWI family proteins MILI and MIWI2 are involved in production of Piwi-interacting RNAs (piRNAs) in fetal male germ cells through a ping-pong amplification cycle. Expression of retrotransposons is elevated in MILI- and MIWI2-deficient male germ cells due to defective de novo DNA methylation, which is presumably caused by impaired piRNA expression. Here, we report that essentially the same abnormalities are observed in MVH (mouse VASA homolog)-deficient mice. Comprehensive analysis of piRNAs in MVH-deficient fetal male germ cells showed that MVH plays crucial roles in the early phase of the ping-pong amplification cycle.
Figures
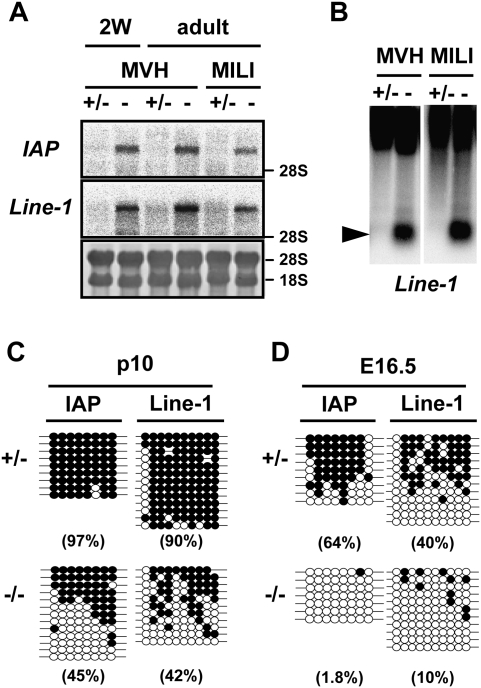
Expression of the IAP and Line-1 retrotransposons and DNA methylation of their regulatory regions in the MVH-deficient testes. (A) Northern blotting analysis of IAP and Line-1 retrotransposons in the testes of 2-wk-old and 2-mo-old MVH- and MILI-deficient mice. The 3′ noncoding region of IAP and the 5′ noncoding regions of Line-1 were used as probes. (B) Methylation-sensitive Southern blotting analysis of the Line-1 promoter region. DNA was extracted from the testes of 2-wk-old heterozygous and homozygous mice. The probe was the same as that used in A. (C,D) Bisulfite sequencing of IAP and Line-1. Germ cells were collected from p10 testis (C) and fetal testis (D) using EGFP as a marker. The LTR region from the IΔI-type IAP in chromosomes 3qD and the 5′ noncoding regions of Line-1 were analyzed. Filled and open circles represent methylated and unmethylated CpGs, respectively. The percentages of methylated CpGs are shown in parentheses.
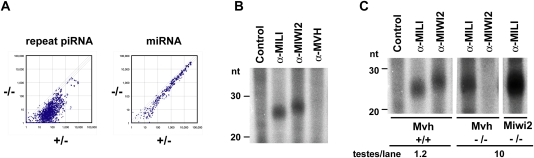
piRNA expression in the MVH-deficient fetal testis. (A) Microarray analysis of the repeat-associated piRNAs and miRNAs. A microarray containing 672 types of repeat-associated piRNA and 150 types of miRNA, both of which were expressed in fetal testes, was produced. Testis RNA samples isolated from E16.5 MVH-deficient and heterozygous mice were used. The values normalized with control probes are plotted, and diagonal lines indicate a 1.5-fold difference in expression. (B) Binding of piRNA with MILI, MIWI2, and MVH. The immunoprecipitated RNAs from E16.5 wild-type testicular lysates were 32P-end-labeled and separated in 15% denaturing urea–polyacrylamide gels. (C) MILI- and MIWI2-bound piRNAs in the mutant embryonic testes. The immunoprecipitated RNAs from E16.5 wild-type and mutant testicular lysates were 32P-end-labeled and separated in 15% denaturing urea–polyacrylamide gels. The numbers of testes used for the immunoprecipitation assays are indicated at the bottom.
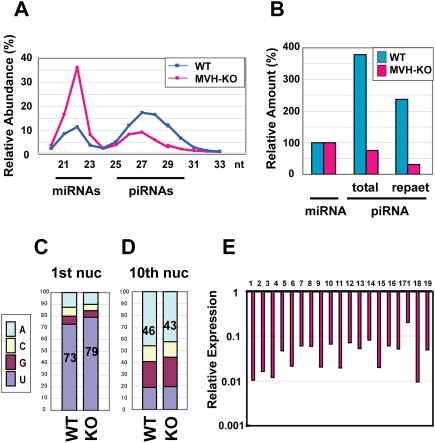
piRNA expression analysis in the MVH-deficient fetal testis by deep sequencing. (A) The size distribution of the total small RNAs in E16.5 testes. Total numbers of small RNAs sequenced from wild-type and MVH-deficient testis libraries were 2,090,035 and 2,812,945, respectively. (B) Ratio of total and repeat-associated piRNA to miRNA. (C,D) Ratio of the first (C) and 10th (D) nucleotides of the repeat-associated piRNAs. (E) Expression of piRNA cluster-derived piRNAs in the MVH-deficient testes. The data of piRNAs derived from the large 19 piRNA clusters are shown. The ratios of piRNAs in the MVH-deficient testes per those in wild-type testes are shown. (Supplemental Table 2) Number 1 represents the data of the largest piRNA cluster, and the following are ordered according to the sizes of the clusters.
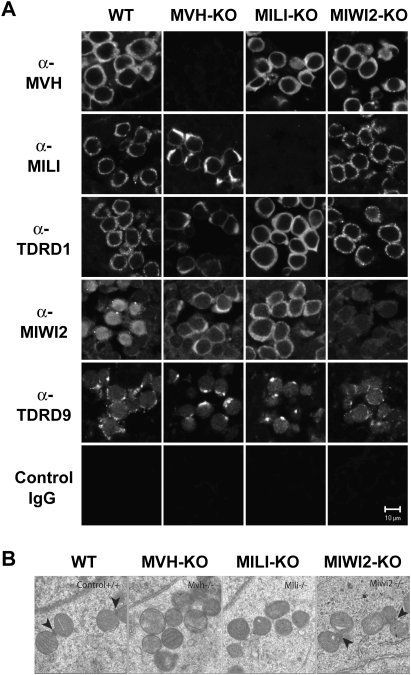
Localization of MILI and MIWI2 in the MVH-deficient fetal germ cells. (A) Immunofluorescence staining of wild-type and mutant testes. Sections of E16.5 testes were stained with anti-MVH, anti-MILI, anti-TDRD1, anti-MIWI2, and anti-TDRD9 antibodies, control IgG. (B) Electron microscopic analyses of MVH-, MILI-, and MIWI2-deficient and control E17.5 testes. Arrowheads indicate intermitochondrial cement.
References
-
- Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ, Kuramochi-Miyagawa S, Nakano T, et al. 2006. A novel class of small RNAs bind to MILI protein in mouse testes. Nature 442: 203–207 - PubMed
-
- Brennecke J, Aravin AA, Stark A, Dus M, Kellis M, Sachidanandam R, Hannon GJ 2007. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 128: 1089–1103 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases