

MicroRNA-340-mediated degradation of microphthalmia-associated transcription factor mRNA is inhibited by the coding region determinant-binding protein
- PMID: 20439467
- PMCID: PMC2898355
- DOI: 10.1074/jbc.M110.109298
MicroRNA-340-mediated degradation of microphthalmia-associated transcription factor mRNA is inhibited by the coding region determinant-binding protein
Retraction in
-
MicroRNA-340-mediated degradation of microphthalmia-associated transcription factor mRNA is inhibited by the coding region determinant-binding protein.J Biol Chem. 2014 Apr 25;289(17):11859. doi: 10.1074/jbc.A110.109298. J Biol Chem. 2014. PMID: 24771639 Free PMC article. No abstract available.
Abstract
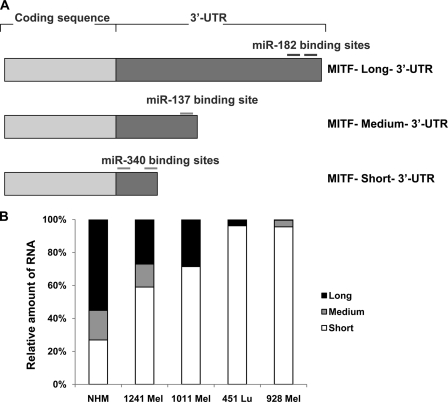
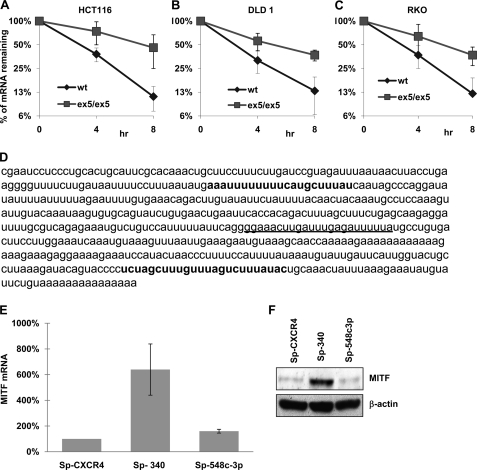
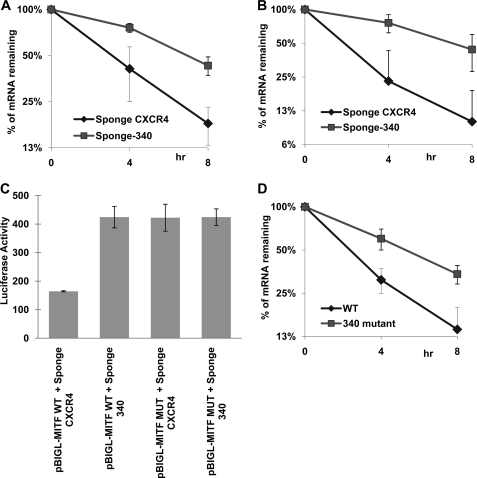
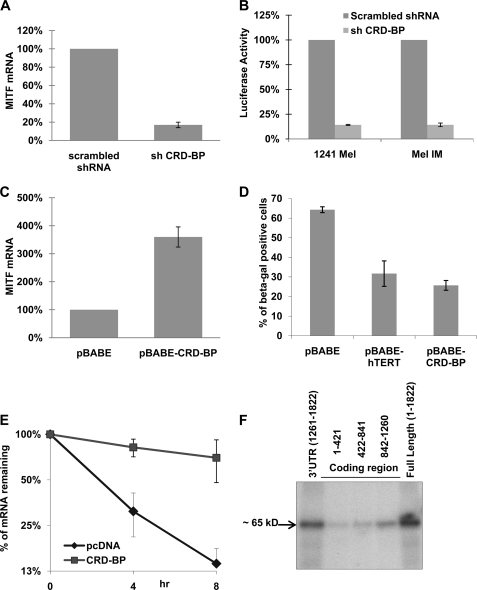
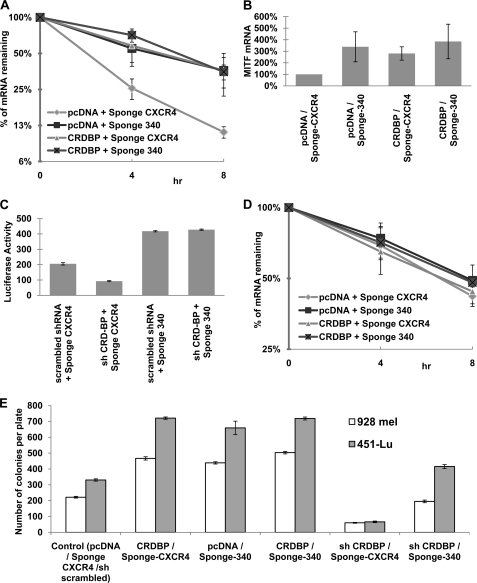
Alternative cleavage and polyadenylation generate multiple transcript variants of mRNA isoforms with different length of 3'-untranslated region (UTR). Alternative cleavage and polyadenylation enable differential post-transcriptional regulation of transcripts via the availability of different cis-acting elements in 3'-UTRs. Microphthalmia-associated transcription factor (MITF) is a master regulator of melanocyte development and melanogenesis. It has also been implicated in melanoma development. Here we show that melanoma cells favor the expression of MITF mRNA with shorter 3'-UTR. This isoform of mRNA is regulated by microRNA, miR-340. miR-340 interacts with two of its target sites on the 3'-UTR of MITF mRNA, causing mRNA degradation and decreased expression and activity of MITF. On the other hand, the RNA-binding protein coding region determinant-binding protein, shown to be highly expressed in melanoma, directly binds to the 3'-UTR of MITF mRNA and prevents the binding of miR-340 to its target sites, resulting in stabilization of the MITF transcript and elevated expression and transcriptional activity of MITF. This interplay between RNA-binding protein and miRNA describes the important mechanism of regulation of MITF in melanocytes and malignant melanomas.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
