

Role of trehalose biosynthesis in Aspergillus fumigatus development, stress response, and virulence
- PMID: 20439478
- PMCID: PMC2897364
- DOI: 10.1128/IAI.00813-09
Role of trehalose biosynthesis in Aspergillus fumigatus development, stress response, and virulence
Abstract
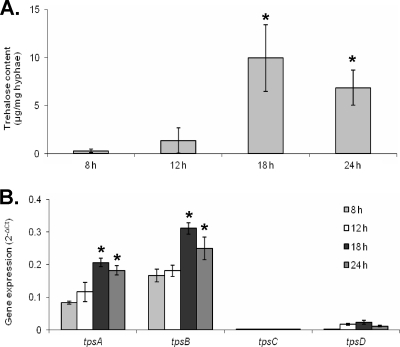
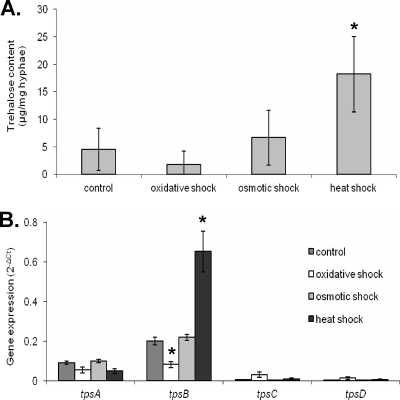
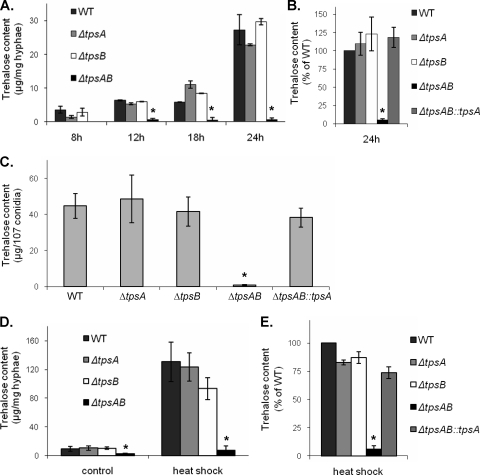
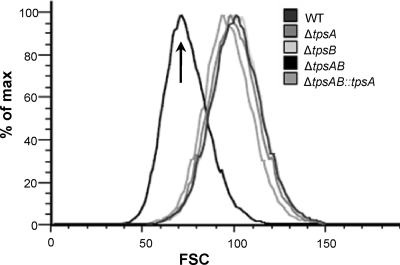
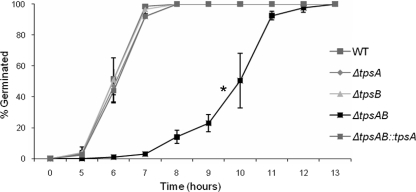
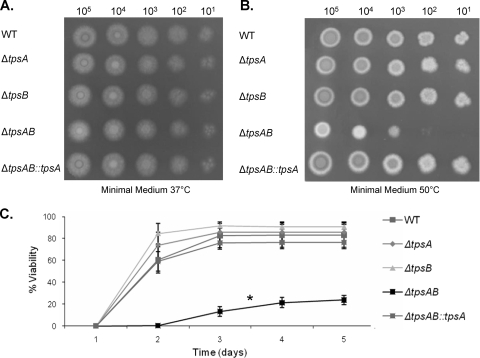
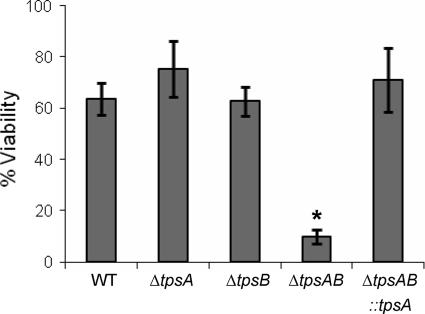
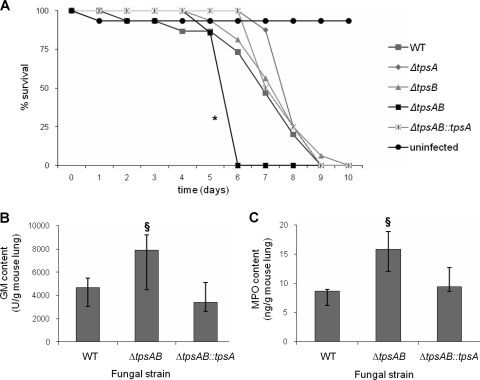
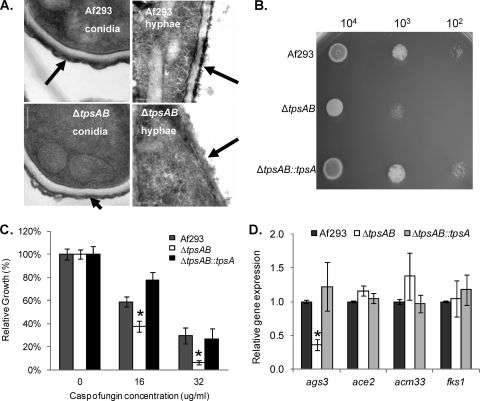
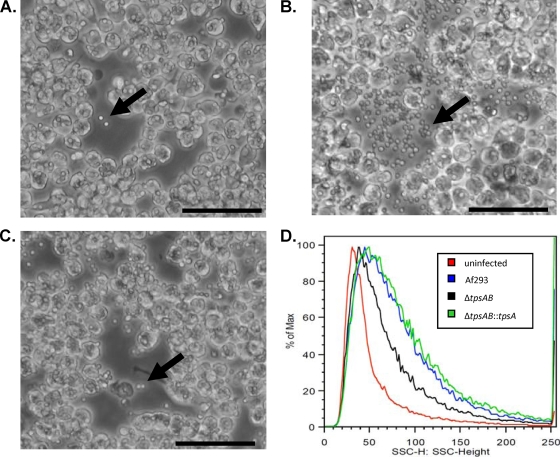
Aspergillus fumigatus is a pathogenic mold which causes invasive, often fatal, pulmonary disease in immunocompromised individuals. Recently, proteins involved in the biosynthesis of trehalose have been linked with virulence in other pathogenic fungi. We found that the trehalose content increased during the developmental life cycle of A. fumigatus, throughout which putative trehalose synthase genes tpsA and tpsB were significantly expressed. The trehalose content of A. fumigatus hyphae also increased after heat shock but not in response to other stressors. This increase in trehalose directly correlated with an increase in expression of tpsB but not tpsA. However, deletion of both tpsA and tpsB was required to block trehalose accumulation during development and heat shock. The DeltatpsAB double mutant had delayed germination at 37 degrees C, suggesting a developmental defect. At 50 degrees C, the majority of DeltatpsAB spores were found to be nonviable, and those that were viable had severely delayed germination, growth, and subsequent sporulation. DeltatpsAB spores were also susceptible to oxidative stress. Surprisingly, the DeltatpsAB double mutant was hypervirulent in a murine model of invasive aspergillosis, and this increased virulence was associated with alterations in the cell wall and resistance to macrophage phagocytosis. Thus, while trehalose biosynthesis is required for a number of biological processes that both promote and inhibit virulence, in A. fumigatus the predominant effect is a reduction in pathogenicity. This finding contrasts sharply with those for other fungi, in which trehalose biosynthesis acts to enhance virulence.
Figures
References
-
- Alvarez-Peral, F. J., O. Zaragoza, Y. Pedreno, and J. C. Arguelles. 2002. Protective role of trehalose during severe oxidative stress caused by hydrogen peroxide and the adaptive oxidative stress response in Candida albicans. Microbiology 148:2599-2606. - PubMed
-
- Beffa, T., F. Staib, J. Lott Fischer, P. F. Lyon, P. Gumowski, O. E. Marfenina, S. Dunoyer-Geindre, F. Georgen, R. Roch-Susuki, L. Gallaz, and J. P. Latge. 1998. Mycological control and surveillance of biological waste and compost. Med. Mycol. 36(Suppl. 1):137-145. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
